
Chromatophore
Saviez-vous ...
Cette sélection se fait pour les écoles par la charité pour enfants en savoir plus . Parrainer un enfant de faire une réelle différence.
Chromatophores sont contenant un pigment réfléchissant la lumière et les cellules présentes dans amphibiens, poissons , reptiles , des crustacés , et céphalopodes. Ils sont en grande partie responsable de la production et de la peau la couleur des yeux dans sang-froid et les animaux sont générés dans le crête neurale cours le développement embryonnaire. Chromatophores matures sont regroupés en sous-classes en fonction de leur couleur (plus exactement " teinte ") sous une lumière blanche: xanthophores (jaune), érythrophores (rouge), iridophores ( réfléchissante / irisé), leucophores (blanc), mélanophores (noir / marron) et cyanophores (bleu). Le terme peut également se référer à, associées à la membrane des vésicules de couleur trouvés dans certaines formes de bactéries photosynthétiques.
Certaines espèces peuvent rapidement changer de couleur par des mécanismes qui translocation pigment et réorienter plaques réfléchissantes dans les chromatophores. Ce procédé est souvent utilisé comme un type de camouflage, est appelé changement de couleur physiologique. Céphalopodes tels que poulpes ont des organes de chromatophores complexes contrôlés par les muscles pour y parvenir, tout en vertébrés tels que caméléons génèrent un effet similaire en la signalisation cellulaire. Ces signaux peuvent être hormones ou neurotransmetteurs et peuvent être initiés par des changements d'humeur, la température, le stress ou des changements visibles dans l'environnement local.
Contrairement animaux à sang froid, les mammifères et les oiseaux ont une seule classe de type de cellule chromatophore comme: la mélanocytes. L'équivalent de sang-froid, mélanophores, sont étudiés par les scientifiques à comprendre les maladies humaines et utilisé comme un outil dans la découverte de médicaments.
Classification
Invertébrés cellules pigmentaires portant ont d'abord été décrits comme chromoforo dans une revue scientifique italienne en 1819. Le terme a été adopté chromatophore tard que le nom de cellules pigmentaires portant dérivées de la crête neurale de sang-froid les vertébrés et les céphalopodes. Le mot lui-même vient du grec mots Khroma (χρωμα) signifie «couleur», et phoros (φορος) signifiant "portant". En revanche, le mot chromatocyte (cyte ou κυτε être grec pour «cellule») a été adopté pour les cellules responsables de la couleur chez les oiseaux et les mammifères. Seul un tel type de cellule, le mélanocyte, a été identifié chez ces animaux.
Ce ne était pas jusqu'à ce que les années 1960 que la structure et la coloration des chromatophores ont compris assez bien pour permettre le développement d'un système de sous-classification basée sur leur apparence. Ce système de classification persiste à ce jour, même si des études plus récentes ont révélé que certains biochimiques aspects des pigments peuvent être plus utiles à une compréhension scientifique de la façon dont fonctionnent les cellules.
Couleur-production tombe en classes distinctes: biochromes, schemochromes . Les véritables biochromes comprennent des pigments, tels que caroténoïdes et ptéridines. Ces pigments absorbent sélectivement des parties de la spectre de la lumière visible qui compose la lumière blanche tout en permettant d'autre longueurs d'onde pour atteindre l'oeil de l'observateur. Schemochromes, également appelés "couleurs", produisent structurelles coloration en réfléchissant des longueurs d'onde (couleurs) et la transmission de la lumière d'autres, en provoquant des ondes lumineuses se immiscer dans la structure ou par diffusion de lumière qui tombe sur eux.
Alors que tous les chromatophores contiennent des pigments ou des structures réfléchissantes (sauf quand il ya eu une génétique mutation conduisant à un trouble comme l'albinisme ), toutes les cellules non pigmentées contenant sont chromatophores. L'hème, par exemple, est un biochrome responsable de l'apparition de sang rouge. Il est principalement trouvé dans globules rouges (érythrocytes), qui sont produits dans la moelle osseuse pendant toute la vie d'un organisme, plutôt que d'être formé au cours du développement embryonnaire. Par conséquent érythrocytes sont pas classés comme chromatophores.

Xanthophores et érythrophores
Chromatophores qui contiennent de grandes quantités de des pigments jaunes de ptéridine sont nommés xanthophores et ceux avec un excès de rouge / caroténoïdes oranges appelés érythrophores. On a découvert que la ptéridine et caroténoïdes contenant vésicules sont parfois trouvés dans la même cellule, et que la couleur globale dépend du rapport de pigments rouges et jaunes. Par conséquent, la distinction entre ces deux types de chromatophores est essentiellement arbitraire. La capacité à générer de ptéridines guanosine triphosphate est une caractéristique commune à la plupart des chromatophores, mais xanthophores semblent avoir voies biochimiques supplémentaires qui se traduisent par une accumulation excessive de pigment jaune. En revanche, les caroténoïdes sont métabolisé de l'alimentation et transportés vers érythrophores. Ce fut d'abord démontré par l'élevage des grenouilles vertes normalement sur un régime de carotène restreinte grillons. L'absence de carotène dans l'alimentation de la grenouille signifiait le rouge / orange caroténoïde couleur «filtre» ne était pas présent dans érythrophores. Il en est résulté la grenouille apparaissant de couleur bleue, au lieu de verte.
Iridophores et leucophores
Iridophores, parfois aussi appelés guanophores, sont des cellules pigmentaires qui reflètent la lumière en utilisant des plaques de chemochromes cristallins fabriqués à partir de guanine. Lorsqu'il est allumé qu'ils génèrent couleurs chatoyantes en raison de la diffraction de la lumière dans les plaques empilées. Orientation de la schemochrome détermine la nature de la couleur observée. En utilisant biochromes que des filtres colorés, iridophores créent un effet optique connu sous le nom Tyndall ou La diffusion de Rayleigh, la production lumineuse bleu ou couleurs vertes.
Un type connexe de chromatophore, le leucophore, se trouve dans certains poissons, en particulier dans le tapetum lucidum. Comme iridophores, ils utilisent cristallin purines (souvent guanine) de réfléchir la lumière. Contrairement iridophores, cependant, ont leucophores cristaux plus organisés qui réduisent diffraction. Compte tenu d'une source de lumière blanche, ils produisent un brillance blanc. Comme xanthophores et érythrophores, dans les poissons la distinction entre iridophores et leucophores ne est pas toujours évident, mais généralement iridophores sont considérés pour générer irisées ou métalliques couleurs tout leucophores produisent des teintes blanches réfléchissantes.
Mélanophores

Mélanophores contiennent eumélanine, un type de la mélanine, qui apparaît noir ou foncé brun en raison de ses qualités absorbant la lumière. Il est conditionné dans des vésicules appelées mélanosomes et répartis dans toute la cellule. L'eumélanine est généré à partir de tyrosine en une série de réactions chimiques catalysées. Ce complexe est un produit chimique contenant des unités de dihydroxyindole-2-dihydroxyindole et l'acide carboxylique avec une certaine cycles pyrrole. L'enzyme clé de la synthèse de mélanine est tyrosinase. Lorsque cette protéine est défectueux, pas de mélanine peut être généré résultant dans certains types d'albinisme. Dans certaines espèces d'amphibiens il ya d'autres pigments emballés aux côtés eumélanine. Par exemple, une nouvelle profondeur pigment de couleur rouge a été identifié dans les mélanophores de phyllomedusine grenouilles . Ce fut par la suite identifié comme pterorhodin, une ptéridine dimère qui se accumule autour de l'eumélanine. Se il est probable que d'autres espèces moins étudiées ont pigments mélanophores complexes, il est néanmoins vrai que la majorité des mélanophores étudié à ce jour ne contiennent exclusivement eumélanine.
Les humains ont une seule classe de cellules pigmentaires, l'équivalent de mammifères mélanophores, pour générer la peau, les cheveux et la couleur des yeux. Pour cette raison, et parce que le grand nombre et la couleur contrastée des cellules font généralement très faciles à visualiser, mélanophores sont de loin le plus largement étudié chromatophore. Cependant, il ya des différences entre la biologie des mélanophores et mélanocytes. En plus de l'eumélanine, les mélanocytes peuvent générer un pigment jaune / rouge appelé phaeomélanine.

Cyanophores
En 1995, il a été démontré que les couleurs bleu éclatant dans certains types de Les poissons mandarins sont pas générés par schemochromes. Au lieu de cela, un biochrome cyan du inconnue nature chimique est responsable. Ce pigment, a trouvé dans les vésicules dans au moins deux espèces de poissons d'callionymid, est très inhabituel dans le animale royaume, comme tous les autres colorants bleus jusqu'à présent étudiés sont schemochromatic. Par conséquent, un nouveau type de chromatophore, le cyanophore, a été proposé. Bien qu'ils semblent inhabituelle dans leur restriction taxonomique, il peut y avoir cyanophores (ainsi que les types de chromatophores plus inhabituelles) dans d'autres poissons et les amphibiens. Par exemple, chromatophores couleurs vives avec des pigments non définies ont été observées à la fois grenouilles venimeuses et grenouilles de verre.
Pigment translocation
De nombreuses espèces ont la capacité de déplacer le pigment à l'intérieur des chromatophores, résultant en un changement visible de la couleur. Ce processus, connu sous le nom changement de couleur physiologique, est le plus largement étudié dans mélanophores, puisque la mélanine est le pigment le plus sombre et le plus visible. Dans la plupart des espèces avec une couche relativement mince derme, les mélanophores dermiques ont tendance à être plat et couvrir une grande surface. Cependant, chez les animaux avec des couches dermiques épais, tels que les reptiles adultes, mélanophores dermiques forment souvent des unités tridimensionnels avec d'autres chromatophores. Ces unités cutanée de chromatophores (DCU) se composent d'un xanthophore supérieure ou erythrophore couche, puis une couche de iridophore, et enfin une couche mélanophore de panier avec des processus couvrant les iridophores.
Les deux types de mélanophores dermiques sont important dans le changement de couleur physiologique. Appartement mélanophores dermiques souvent superposer autres chromatophores alors quand le pigment est dispersé dans toute la cellule de la peau apparaît sombre. Lorsque le pigment est agrégé en direction du centre de la cellule, les pigments dans d'autres chromatophores sont exposés à la lumière et la peau prend leur teinte. De même, après l'agrégation de mélanine dans DCU, la peau apparaît en vert grâce xanthophore (jaune) le filtrage de la lumière diffusée à partir de la couche de iridophore. Sur la dispersion de la mélanine, la lumière ne est plus diffusée et la peau apparaît sombre. Comme les autres chomatophores biochromatic sont également capables de translocation de pigment, les animaux avec des types de chromatophores multiples peuvent générer une gamme spectaculaire de couleurs de peau en faisant bon usage de l'effet de la division.,

Le contrôle et la mécanique de la translocation de pigment rapide a été bien étudié dans un certain nombre de différentes espèces, notamment les amphibiens et les téléostéens poissons., Il a été démontré que le processus peut être sous hormonal , contrôle neuronal ou les deux. Neurochimiques connues pour déplacer pigment comprennent noradrénaline, par son récepteur sur la surface de mélanophores. Les hormones primaires impliqués dans la régulation translocation semblent être les mélanocortines, la mélatonine et Hormone concentrant mélanine (MCH), qui sont produites principalement dans l'hypophyse, glande pinéale et l'hypothalamus, respectivement. Ces hormones peuvent également être générés dans un paracrine par les cellules de la peau. A la surface de la mélanophores a été démontré que les hormones pour activer spécifique Les récepteurs aux protéines G qui, à son tour, la transduction du signal dans la cellule couplée. Mélanocortine conduisent à la dispersion de pigment, tandis que la mélatonine et les résultats de SMI dans l'agrégation.
Mélanocortine nombreux, SMI et récepteurs de la mélatonine ont été identifiées dans les poissons et les grenouilles, y compris une homologue de MC1R, un récepteur de la mélanocortine connu pour réguler peau et la couleur des cheveux chez l'homme. A l'intérieur de la cellule, l'adénosine monophosphate cyclique (cAMP) a été montré pour être un élément important second messager de translocation de pigment. Grâce à un mécanisme non encore entièrement compris, les influences d'AMPc d'autres protéines telles que la protéine kinase A à conduire moteurs moléculaires portant pigments contenant des vésicules le long des deux microtubules et microfilaments.,,
adaptation d'arrière-plan
La plupart des poissons, reptiles et amphibiens subissent un changement de couleur physiologique limitée en réponse à un changement dans l'environnement. Ce type de camouflage, connu comme arrière-plan d'adaptation, apparaît le plus souvent comme un léger assombrissement ou un éclaircissement du teint de la peau à environ imiter la teinte de l'environnement immédiat. Il a été démontré que le processus d'adaptation de fond dépend vision (il semble que l'animal doit être en mesure de voir l'environnement pour s'y adapter), et que la mélanine translocation dans mélanophores est le principal facteur de changement de couleur. Certains animaux, comme les caméléons et anoles, ont une réponse d'adaptation de fond fortement développé capable de générer un certain nombre de différentes couleurs très rapidement. Ils ont adapté la capacité à changer de couleur en fonction de la température, de l'humeur, le niveau de stress et les signaux sociaux, plutôt que de simplement imiter leur environnement.

Développement
Pendant vertébrés le développement embryonnaire, sont les chromatophores une d'un certain nombre de types de cellules générées dans le crête neurale, une bande appariée de cellules résultant en marge de la tube neural. Ces cellules ont la capacité de migrer sur de longues distances, ce qui permet aux chromatophores peuplent de nombreux organes du corps, y compris la peau, les yeux, les oreilles et le cerveau. Laissant la crête neurale dans les vagues, chromatophores prennent soit une route dorsolatéral à travers le derme, entrant dans la ectoderme à travers de petits trous dans la lame basale, ou un itinéraire entre le ventromédian somites et le tube neural. L'exception à cette règle les mélanophores de l'épithélium pigmentaire de la rétine de l'œil. Ce ne sont pas dérivées de la crête neurale, au lieu d'une évagination du tube neural génère le cupule optique qui, à son tour, forme la rétine .
Quand et comment cellules précurseurs de chromatophore multipotentes (appelés chromatoblasts) développer dans leurs sous-types de fille est un domaine de recherche en cours. Il est connu dans les embryons de poisson zèbre, par exemple, que par les 3 jours après fécondation chacune des classes présentes dans le poisson adulte de cellules - mélanophores, xanthophores et iridophores - sont déjà présents. Des études utilisant des poissons mutants ont démontré que les facteurs de transcription tels que kit, SOX10 et MITF sont importantes dans le contrôle de la différenciation des chromatophore. Si ces protéines sont défectueux, chromatophores peuvent être régional ou totalement absent, résultant en une leucistic trouble.
Les applications pratiques
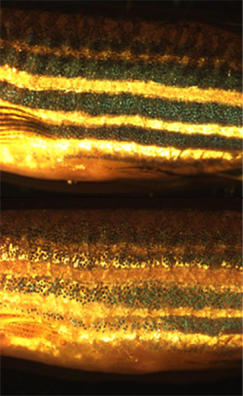
En plus de la recherche fondamentale sur une meilleure compréhension des chromatophores eux-mêmes, les cellules sont utilisées à des fins de recherche appliquée. Par exemple, larves de poisson zèbre sont utilisés pour étudier comment chromatophores organiser et de communiquer pour générer précision le motif à rayures horizontales régulière comme on le voit dans les poissons adultes. Ceci est considéré comme une utile système modèle pour comprendre la structuration dans le évolutif champ de la biologie du développement. Chromatophore biologie a également été utilisée pour modéliser l'état ou de la maladie humaine, y compris mélanome et l'albinisme. Récemment, le gène responsable de la souche spécifique de poisson zèbre d'or mélanophores, SLC24A5, se est trouvé avoir un équivalent humain qui est fortement corrélé avec couleur de la peau.
Chromatophores sont également utilisés en tant que biomarqueur de cécité chez les espèces à sang froid, comme des animaux avec certains défauts visuels ne parviennent pas à fond adapter à des environnements lumineux. Des homologues humains des récepteurs qui médient pigment translocation dans les mélanophores sont considérés comme impliqués dans des processus tels que suppression de l'appétit et bronzage, ce qui en fait des cibles attrayantes pour médicaments. Pourquoi les entreprises pharmaceutiques ont développé une dosage biologique pour identifier rapidement des composés bioactifs potentiels en utilisant des mélanophores de la grenouille africaine à griffes . D'autres scientifiques ont développé des techniques pour l'utilisation de mélanophores que biocapteurs, et pour la détection rapide de la maladie (sur la base sur la découverte que blocs de toxine pertussis d'agrégation de pigment en mélanophores de poisson). Potentiel applications militaires de chromatophores médiation changements de couleur ont été proposées, principalement comme un type de camouflage actif.
Chromatophores céphalopodes

Céphalopodes ont Coleoid complexes multicellulaires «organes» qu'ils utilisent pour changer rapidement de couleur. Ce est le plus notable dans couleurs vives calmar, seiches et pieuvres. Chaque unité de chromatophore est composé d'une cellule de chromatophore unique et de nombreux muscles, des nerfs, gliales et cellules gaine. A l'intérieur de la cellule de chromatophore, en granulés pigmentaires sont enfermés dans un sac élastique, dite saccule cytoelastic. Pour changer la couleur de l'animal déforme la forme ou la taille de sacculus par la contraction musculaire, en changeant sa translucidité, la réflectivité ou opacité. Ceci diffère du mécanisme utilisé dans les poissons, les amphibiens et les reptiles, en ce que la forme de la saccule est modifiée plutôt que d'une translocation des vésicules de pigment dans la cellule. Cependant un effet similaire est obtenu.
Poulpes opèrent dans chromatophores, affiche chromatiques ondulatoires complexes, résultant dans une variété de l'évolution rapide des schémas de couleurs. Les nerfs qui exploitent les chromatophores sont pensés pour être positionné dans le cerveau, dans un ordre semblable à les chromatophores ils chaque contrôle. Cela signifie que le modèle de changement de couleur correspond au modèle d' activation neuronale . Cela peut expliquer pourquoi, que les neurones sont activés l'un après l'autre, le changement de couleur se produit dans les vagues. Comme les caméléons, les céphalopodes utilisent changement de couleur physiologique pour l'interaction sociale. Ils sont aussi parmi les plus compétents au fond adaptation, ayant la capacité de faire correspondre la couleur et la texture de leur environnement local avec une précision remarquable.
Bactéries
Chromatophores sont également présents dans les membranes de bactéries phototrophes. Utilisé principalement pour la photosynthèse, elles contiennent pigments caroténoïdes et bactériochlorophylle. En bactéries pourpres, tels que Rhodospirillum rubrum les protéines collecteurs de lumière sont intrinsèques aux membranes de chromatophores. Toutefois, Chlorobi ils sont disposés dans spécialisé les complexes d'antenne appelés chlorosomes.