
Parasaurolophus
À propos de ce écoles sélection Wikipedia
Cette sélection Wikipedia est déconnecté disponibles à partir enfants SOS pour la distribution dans le monde en développement. Avec enfants SOS vous pouvez choisir de parrainer des enfants dans plus de cent pays
| Parasaurolophus Plage temporelle: Crétacé supérieur, 76.5-73Ma | |
|---|---|
 | |
| P. cyrtocristatus, Field Museum of Natural History | |
| Classification scientifique | |
| Uni: | Animalia |
| Embranchement: | Chordata |
| Classe: | Reptilia |
| Super: | Dinosauria |
| Ordre: | † Ornithischia |
| Famille: | † Hadrosauridae |
| Sous-famille: | † Lambeosaurinae |
| Genre: | † Parasaurolophus Parcs, 1922 |
| Espèce | |
| |
Parasaurolophus (pron .: / ˌ p ær ə s ɔː r ɒ l ə fa ə s / PARR -ə-scie ROL -ə-fəs ou / ˌ p ær ə ˌ s ɔr ə l oʊ fa ə s / PARR -ə- Sawr de LOH -fəs; ce qui signifie «lézard près aigrettes» en référence à Saurolophus) est un genre de ornithopode dinosaure qui vivait dans ce qui est maintenant l'Amérique du Nord au cours de la Crétacé supérieur Période, il ya environ 76,5 à 73.000.000 années. Ce était un herbivore qui marchait à la fois comme bipède et un quadrupède. Trois espèces sont reconnues: P. walkeri (le tapez espèces), P. tubicen et le P. court aigrettes cyrtocristatus. Vestiges sont connus à partir Alberta ( Canada ), et Nouveau-Mexique et Utah ( États-Unis ). Le genre a été décrite pour la première en 1922 par William Parcs à partir d'un crâne et squelette partiel en Alberta.
Parasaurolophus était hadrosaurid, partie d'une diversité famille de dinosaures du Crétacé connus pour leur gamme de parures de tête bizarres. Ce genre est connu pour sa grande crête crânienne élaboré, qui, à ses plus grandes formes d'un long tube incurvé saillie vers le haut et l'arrière de la crâne. Charonosaurus de la Chine , qui peut avoir été son plus proche parent, avait un crâne semblable et potentiellement une crête similaire. La crête a été beaucoup discutée par les scientifiques; le consensus est que les principales fonctions comprenaient la reconnaissance visuelle des deux espèces et le sexe, la résonance acoustique, et thermorégulation. Il est l'un des rares hadrosaurids, connus à partir de seulement une poignée de bons spécimens.
Description

Comme la plupart des dinosaures, le squelette de Parasaurolophus est incomplètement connue. La longueur de la tapez spécimen de P. walkeri est estimé à 9,5 m (31 pi), et son poids est estimé à 2,5 tonnes (2,8 tonnes courtes). Son crâne est environ 1,6 m (5 pi 3 po) de longueur, y compris la crête, alors que le crâne de type de P. tubicen est plus de 2 m (6 pi 7 po) de long, indiquant un plus grand animal. Son seul membre antérieur connu a été relativement courte pour un hadrosaurid, avec un court mais large omoplate. Le fémur mesure 103 cm (41 po) de long dans P. walkeri est robuste et de sa longueur par rapport aux autres hadrosaurids. Le bras et os du bassin ont été également fortement construites.
Comme d'autres hadrosaurids, il était capable de marcher sur deux jambes soit ou quatre. Il a probablement préféré chercher de la nourriture à quatre pattes, mais a couru sur deux. Le épines neurales de la vertèbres étaient grands, comme cela était courant dans lambeosaurines; plus haut sur les hanches, ils ont augmenté la hauteur du dossier. impressions de la peau sont connus pour P. walkeri, montrant échelles tuberculeux comme uniformes mais pas de grandes structures.

La caractéristique la plus notable était la crête crânienne, qui sortait de l'arrière de la tête et a été constitué de la prémaxillaire et os du nez. Le P. Type walkeri spécimen a une encoche dans les épines neurales près du lieu où la crête aurait frappé le dos, mais cela peut être une pathologie particulière à cet individu. William Parks, qui a nommé le genre, l'hypothèse qu'un ligament couru de la crête à l'encoche pour soutenir la tête. Bien que cette idée semble peu probable, Parasaurolophus est parfois restauré avec un lambeau de peau de la crête au cou. La crête était creux, avec des tubes distincts menant de chaque narine à la fin de la crête avant de renverser la direction et de redescendre la crête et dans le crâne. Les tubes ont été simples dans P. walkeri et plus complexe dans P. tubicen, où certains tubes étaient aveugles et d'autres se sont réunis et séparés. Alors que P. walkeri et P. tubicen avait de longues crêtes avec seulement une légère courbure, P. cyrtocristatus avait une courte crête avec un profil plus circulaire.
Classification
Comme son nom l'indique, Parasaurolophus a d'abord pensé être étroitement liée à Saurolophus en raison de sa crête superficiellement similaire. Toutefois, il a rapidement été réévalué en tant que membre de la lambeosaurine sous-famille des hadrosaurids- Saurolophus est un hadrosaurine. Il est généralement interprétée comme une ramification séparée des lambeosaurines, distinct du casque à aigrettes Corythosaurus, Hypacrosaurus, et Lambeosaurus. Son plus proche parent connu semble être Charonosaurus, un lambeosaurine avec un crâne similaire (mais pas encore complet crête) de l' Amur région du nord de la Chine, et les deux peuvent former un clade Parasaurolophini. P. cyrtocristatus, avec son court, rond crête, peut être le plus basal des trois espèces de Parasaurolophus connus, ou il peut représenter spécimens subadultes ou femelles de P. tubicen.
Découverte et nommage

Signification «lézard à crête près", le nom Parasaurolophus est dérivé du grec para / παρα "à côté" ou, saurus / σαυρος "lézard" "proche" et lophos / λοφος "crête". C'est basé sur ROM 768, un crâne et le squelette partielle manquant la plupart de la queue et les pattes arrières-dessous des genoux, qui a été trouvé par une partie du champ de la Université de Toronto en 1920 près de Sand Creek le long de la La rivière Red Deer en Alberta, Canada. Ces roches sont maintenant connus comme la Campanien-âge Crétacé supérieur Formation de Dinosaur Park. William Parcs nommé le spécimen P. walkeri en l'honneur de Sir Byron Edmund Walker, président du conseil d'administration de la Musée royal de l'Ontario. Parasaurolophus vestiges sont rares en Alberta, avec une seule autre crâne partiel du (probablement) la Formation Dinosaur Park, et trois spécimens de Dinosaur Park manquant crânes, pouvant appartenir au genre. Dans certaines listes fauniques, il est fait mention d'une éventuelle P. dans le matériau walkeri Formation de Hell Creek Montana, une unité de roche Maastrichtien tard. Cet événement ne est pas noté par Sullivan et Williamson dans leur examen du genre 1999, et n'a pas été précisé lors de ailleurs.
En 1921, Charles H. Sternberg a récupéré un crâne partiel ( PMU.R1250) de ce qui est maintenant connu sous le nom légèrement plus jeune Formation Kirtland dans Comté de San Juan, au Nouveau-Mexique. Ce spécimen a été envoyé à Uppsala, Suède , où Carl Wiman a décrit comme une seconde espèces, P. tubicen, en 1931. L'épithète spécifique est dérivé du latin tǔbǐcěn "trompettiste". Une seconde, presque complet P. tubicen crâne ( NMMNH P-25100) a été trouvé dans le Nouveau-Mexique en 1995. Utilisation tomodensitométrie de ce crâne, Robert Sullivan et Thomas Williamson a donné un genre traitement monographique en 1999, couvrant les aspects de son anatomie et de la taxonomie, et les fonctions de sa crête. Williamson publié plus tard un examen indépendant de la dépouille, en désaccord avec les conclusions taxonomiques.
John Ostrom décrit un autre bon spécimen ( FMNH P27393) du Nouveau-Mexique que P. cyrtocristatus en 1961. Il comprend un crâne partiel avec une courte, crête arrondie, et la plupart des squelette postcrânien sauf pour les pieds, le cou, et des parties de la queue. Son nom spécifique est dérivé du latin curtus "raccourci" et cristatus "huppé". Le spécimen a été trouvé dans sa partie supérieure de la Formation Fruitland ou, plus probablement, la base de la formation sus-jacente Kirtland. La gamme de cette espèce a été élargi en 1979, quand David B. et Weishampel James A. Jensen a décrit un crâne partiel avec une crête similaire ( BYU 2467) dans le Campanien-âge Formation des Kaiparowits Garfield County, Utah. Depuis lors, un autre crâne a été trouvé dans l'Utah avec le court / tour P. cyrtocristatus crête morphologie.
Espèce

Le espèce type P. walkeri, de l'Alberta, est connu par un seul spécimen définitives de la Dinosaur Park Formation, bien que d'autres de la Dinosaur Park appartient probablement à elle. Elle diffère de P. tubicen en ayant tubes simples dans sa crête, et de P. cyrtocristatus en ayant une longue crête non arrondie et un plus bras de avant-bras. Il a vécu il ya entre 76,5 et 75,3 millions années.
P. tubicen, du Nouveau-Mexique, est connu à partir des restes d'au moins trois personnes. Ce est la plus grande espèce, avec plus de passages d'air complexes dans sa crête que P. walkeri, et une crête plus droite que P. cyrtocristatus. P. tubicen ne est connue que de la De-na-zin membres de la Formation Kirtland, et a vécu il ya environ 73,4 à 73.000.000 années, ce qui rend les espèces plus récentes.
P. cyrtocristatus, à partir de la Kaiparowits et Fruitland Formations du Nouveau-Mexique et l'Utah, est connu à partir de trois échantillons possibles. Il a vécu il ya 75,5 à 74.500.000 années, et est la plus petite espèce, avec une crête arrondie courte. Sa petite taille et la forme de sa crête ont conduit plusieurs scientifiques à suggérer qu'il représente mineurs ou femelles de P. tubicen, si P. tubicen vécu au moins un million d'années après. Comme l'a noté Thomas Williamson, le matériau de type P. cyrtocristatus est d'environ 72% la taille de P. tubicen, près de la taille à laquelle d'autres lambeosaurines sont interprétés pour commencer montrant définitif dimorphisme sexuel dans leurs crêtes (~ 70% de la taille adulte). Cependant, cette position a été rejetée dans des études récentes de lambeosaurines.
Paléoécologie
Parasaurolophus walkeri, de la Formation de Dinosaur Park, était membre d'un diversifiée et bien documenté la faune d'animaux préhistoriques, y compris les dinosaures bien connus tels que l' cornes Centrosaurus et Chasmosaurus; autres duckbills Gryposaurus et Corythosaurus; tyrannosaurid Gorgosaurus; et blindé Edmontonia, Euoplocephalus et Anodontosaurus. Ce était un constituant rare de cette faune. La Formation de Dinosaur Park est interprété comme un réglage bas-relief des rivières et plaines inondables qui sont devenus plus marécageux et influencé par marins conditions au fil du temps que le Voie maritime intérieure de l'Ouest transgressé vers l'ouest. Le climat était plus chaud aujourd'hui l'Alberta, sans gel, mais avec des saisons humides et sèches. Conifères étaient apparemment la dominante plantes couvert, avec une sous-étage de fougères , fougères arborescentes, et angiospermes.
Certains des hadrosaures moins communs dans la Formation de Dinosaur Park du parc provincial Dinosaur, comme Parasaurolophus, peuvent représenter les restes de personnes décédées lors de la migration à travers la région. Ils pourraient également avoir eu un habitat plus des hautes terres où ils ont peut-être imbriquée ou nourris. La présence de Parasaurolophus et Kritosaurus dans les sites de fossiles de latitude nord peut représenter échanges fauniques entre biomes contraire distinctes nord et sud dans Crétacé supérieur en Amérique du Nord. Les deux taxons sont rares en dehors du biome sud, où, avec Pentaceratops, ils sont prédominent membres de la faune.
Au Nouveau-Mexique et l'Utah, l'espèce P. cyrtocristatus partagé son environnement avec les cornes cératopsiens Utahceratops, Kosmoceratops, et Pentaceratops, et la coelurosaur Ornithomimus et le tyrannosaure Teratophoneus.
La dernière et la plus grande des espèces Parasaurolophus, P. tubicen, a vécu au Nouveau-Mexique aux côtés de la grande sauropode Alamosaurus , bec de canard Kritosaurus, Pentaceratops cornes, blindés Nodocephalosaurus, Saurornitholestes, et la tyrannosaurid Bistahieversor. La Formation de Kirtland est interprété comme plaines inondables apparaissant après une retraite de la Voie maritime intérieure de l'Ouest. Les conifères sont les plantes dominantes et chasmosauriné cornes dinosaures étaient apparemment plus commun que hadrosaurids.
Paléobiologie
Alimentation
En hadrosaurid, Parasaurolophus était un grand herbivore bipède / quadrupède, manger des plantes avec un crâne sophistiquée qui a permis un mouvement de broyage analogue à la mastication. Son dents étaient continuellement remplacées; ils ont été emballés dans des batteries dentaires contenant des centaines de dents, seulement une poignée de qui étaient en cours d'utilisation à tout moment. Il a utilisé son bec au matériel végétal de culture, qui se est tenue dans les mâchoires par un joue comme organe. La végétation aurait pu être prise depuis le sol jusqu'à une hauteur d'environ 4 mètres (13 pieds). Comme l'a noté Bob Bakker, lambeosaurines ont des becs plus étroites que hadrosaurines, ce qui implique que Parasaurolophus et ses parents pourrait nourrir plus sélective que leurs homologues sans crête large bec.
Crête crânienne

De nombreuses hypothèses ont été avancées quant à ce qui fonctionne de la crête crânienne de Parasaurolophus effectuées, mais la plupart ont été discréditées. Il croit maintenant qu'il peut avoir eu plusieurs fonctions: affichage visuel pour identifier les espèces et le sexe, amplification du son pour la communication et la thermorégulation. Il ne est pas clair ce qui était le plus important à quel moment dans l'évolution de la crête et ses voies nasales internes.
Les différences entre les espèces et les stades de croissance
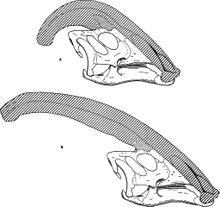
Comme pour les autres lambeosaurines, on pense que la crête crânienne de Parasaurolophus changé avec l'âge et est un caractéristique dimorphisme sexuel chez les adultes. James Hopson, l'un des premiers chercheurs à décrire crêtes de lambeosaurine en termes de telles distinctions, a suggéré que P. cyrtocristatus, avec sa petite crête, était la forme féminine de P. tubicen. Thomas Williamson a suggéré que ce était la forme juvénile. Aucune des deux hypothèses est devenu largement accepté. Comme seulement six bonnes crânes et une boîte crânienne pour mineurs soient connus, du matériel supplémentaire qui contribuera à dissiper ces relations potentielles. Williamson a noté que dans tous les cas, juvénile Parasaurolophus avait probablement petites crêtes arrondies comme P. cyrtocristatus, qui a probablement augmenté plus rapidement que les individus approchés la maturité sexuelle. Réétudier récente d'une boîte crânienne juvénile précédemment affectée à Lambeosaurus, désormais assigné à Parasaurolophus, fournit la preuve que d'une petite crête tubulaire était présent chez les juvéniles. Ce spécimen conserve une petite hausse du torchage os frontaux qui était similaire mais plus petit que ce qui est vu de spécimens adultes; chez les adultes, les frontaux formé une plate-forme qui a soutenu la base de la crête. Ce spécimen indique également que la croissance de la crête de Parasaurolophus et le profil du visage d'individus juvéniles différaient de la Corythosaurus - Hypacrosaurus - modèle Lambeosaurus, en partie parce que la crête de Parasaurolophus n'a pas la osseuse mince »de fat 'qui constitue la partie supérieure du la crête des trois autres lambeosaurines.
Hypothèses rejetées sur la fonction
De nombreuses suggestions rapides axés sur les adaptations pour un mode de vie aquatique, à la suite l'hypothèse que hadrosaurids étaient amphibie, une ligne commune de la pensée jusqu'à ce que les années 1960. Ainsi, Alfred Sherwood Romer a proposé il a servi de tuba, Martin Wilfarth que ce était une pièce jointe pour un mobile trompe utilisés comme un tube de respiration ou pour la cueillette de nourriture, Charles M. Sternberg qu'il a servi comme airtrap pour garder l'eau hors de la les poumons, et Ned Colbert qu'il a servi comme un réservoir d'air pour des séjours prolongés sous-marins.
D'autres propositions sont plus de nature physique. Comme mentionné ci-dessus, William Parks a suggéré qu'il était jointe à la vertèbre avec ligaments ou des muscles, et aidé à déplacer et à soutenir la tête. Othenio Abel proposé il a été utilisé comme une arme dans le combat entre les membres de la même espèce, et Andrew Milner a suggéré qu'il pourrait être utilisé comme un déflecteur de feuillage, comme la crête du casque (appelé 'casque') du casoar . D'autres propositions faites logement organes spécialisés de la fonction majeure. Halszka Osmólska suggéré qu'il occupe glandes à sel, et John Ostrom suggéré qu'il logés zones élargi pour tissu olfactif et beaucoup améliorées sens de l'odorat des lambeosaurines, qui ne avaient pas les capacités défensives évidentes. Une suggestion inhabituelle, faite par créationniste Duane Gish, ce est que la crête logé glandes chimiques qui lui ont permis de jeter des jets de «feu» chimique sur les ennemis, semblable à la moderne-jour scarabée bombardier.
La plupart de ces hypothèses ont été discréditées ou rejetée. Par exemple, il n'y a pas de trou à la fin de la crête pour une fonction de la plongée en apnée. Il n'y a pas de cicatrices musculaires pour une trompe et il est douteux que l'animal avec un bec aurait besoin d'un. Comme un sas proposé, il ne aurait pas gardé hors de l'eau. Le réservoir d'air proposé aurait été insuffisant pour un animal de la taille de Parasaurolophus. Autres hadrosaurids eu de grandes têtes sans avoir besoin de grandes crêtes creux pour servir de points de fixation pour soutenir les ligaments. En outre, aucune des propositions expliquer pourquoi la crête a une telle forme, pourquoi les autres devraient avoir lambeosaurines crêtes qui ressemblent très différent, mais remplir une fonction analogue, comment hadrosaurids sans crête ou solides à aigrettes se entendaient sans ces capacités, ou pourquoi certains hadrosaurids eu solide crêtes. Ces considérations impact particulièrement hypothèses fondées sur l'augmentation des capacités des systèmes déjà présents dans l'animal, comme la glande de sel et olfaction hypothèses, et indiquent que ce ne étaient pas les fonctions primaires de la crête. En outre, les travaux de la cavité nasale de lambeosaurines montre que nerfs olfactifs et tissu sensoriel correspondant étaient en grande partie en dehors de la partie des voies nasales à la crête, de sorte que l'expansion de la crête avait peu à voir avec le sens de l'odorat.
Fonctions sociales

Au lieu de cela, sociale et fonctions physiologiques se sont pris en charge comme fonction (s) de la crête, se concentrant sur et visuelle identification auditive et de communication. Comme un grand objet, la crête a une valeur clairement comme un signal visuel, et définit cet animal en dehors de ses contemporains. La grande taille de hadrosauridé orbites et la présence de anneaux sclérosés dans les yeux impliquent vision aiguë et habitudes diurnes, la preuve que la vue était important de ces animaux. Si, comme ce est souvent illustré, un volant de la peau se étendait de la crête du cou ou du dos, l'affichage visuel proposé aurait été encore plus voyante. Comme cela est suggéré par d'autres crânes lambeosaurine, la crête de Parasaurolophus susceptibles permis à la fois l'identification des espèces (comme le séparant de Corythosaurus ou Lambeosaurus) et l'identification sexuelle par la forme et la taille.
Fonction Sounding
Cependant, l'aspect extérieur de la crête ne correspond pas à l'anatomie interne complexe des voies nasales, ce qui suggère une autre fonction a représenté l'utilisation de l'espace intérieur. Carl Wiman a été le premier à proposer, en 1931, que les passages servi une fonction de signalisation auditive, comme un crumhorn; Hopson et David B. Weishampel revisité cette idée dans les années 1970 et 1980. Hopson trouvé qu'il existe des preuves anatomique qui avaient hadrosaurids audience forte. Il existe au moins un exemple, dans le Corythosaurus est associée, d'une mince étrier (reptilienne os de l'oreille) en place, qui, combiné avec un grand espace pour un tympan implique une sensible oreille moyenne. En outre, le hadrosaurid lagena est allongé comme un de crocodiliens, indiquant que la partie auditive du l'oreille interne a été bien développé. Weishampel suggéré que P. walkeri était capable de produire des fréquences de 48 à 240 Hz, et P. cyrtocristatus (interprétée comme une forme de crête juvénile) de 75 à 375 Hz. Basé sur la similitude des hadrosaurid oreilles internes à celles des crocodiles , il a également proposé que hadrosaurids adultes sont sensibles aux hautes fréquences, tels que leur progéniture pourrait produire. Selon Weishampel, cela est compatible avec les parents et les enfants à communiquer.
La modélisation informatique d'un spécimen bien conservés de P. tubicen, avec des passages d'air plus complexes que ceux de P. walkeri, a permis la reconstruction du possible sonner sa crête produite. Le chemin principal résonne à environ 30 Hz, mais le sinus anatomie compliquée et provoque des pics et des vallées dans le son.
Fonction de refroidissement
La grande surface de contact et vascularisation de la crête suggère également une fonction de thermorégulation. PE Wheeler premier suggéré cette utilisation en 1978 comme un moyen de garder le cerveau cool. Teresa Maryańska et Osmólska également proposé thermorégulation à peu près au même moment, et Sullivan et Williamson ont plus d'intérêt. David Evans de discussion des fonctions de crête lambeosaurine 2006 a été favorable à l'idée, au moins comme un facteur initial pour l'évolution de l'expansion de crête.