
Albertosaurus
Saviez-vous ...
Cette sélection Wikipedia est déconnecté disponibles à partir enfants SOS pour la distribution dans le monde en développement. SOS Enfants a regardé des enfants en Afrique depuis quarante ans. Pouvez-vous aider dans leur travail en Afrique ?
| Albertosaurus sarcophagus Plage temporelle: Crétacé supérieur, 71-68Ma | |
|---|---|
 | |
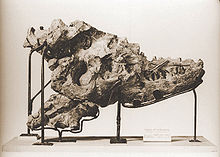
| Crâne et le cou d'un moulage du squelette monté, Muséum national d'histoire naturelle | |
| Classification scientifique | |
| Uni: | Animalia |
| Embranchement: | Chordata |
| Classe: | Reptilia |
| Ordre: | Saurischia |
| Famille: | † Tyrannosauridae |
| Sous-famille: | † Albertosaurinae |
| Genre: | † Albertosaurus Osborn, 1905 |
| Espèce: | † A. sarcophage |
| Nom binomial | |
| Albertosaurus sarcophagus Osborn, 1905 | |
| Synonymes | |
Deinodon sarcophage (Osborn, 1905) | |
Albertosaurus (pron .: / æ l ˌ b ɜr t ɵ s ɔr ə s /; ce qui signifie «lézard Alberta») est un genre de tyrannosaurid théropode dinosaure qui a vécu dans l'ouest de l'Amérique du Nord au cours de la fin du Crétacé Période, environ 70 il ya des millions d'années. Le tapez espèces, A. sarcophage, était apparemment restreint plage au moderne-jour Canada province de Alberta, après quoi le genre est nommé. Les scientifiques ne se entendent pas sur le contenu du genre, avec une certaine reconnaissance Gorgosaurus libratus comme une deuxième espèce.
En tyrannosaurid, Albertosaurus était un bipède prédateur avec les mains minuscules, deux-doigts et une tête massive avec des dizaines de grandes dents pointues. Il peut avoir été au sommet de la chaîne alimentaire dans son locale écosystème. Bien que relativement grand pour un théropode, Albertosaurus était beaucoup plus petit que son célèbre rapport Tyrannosaurus , probablement un poids inférieur à 2 tonnes métriques.
Depuis la première découverte en 1884, des fossiles de plus de trente personnes ont été récupérés, fournissant aux scientifiques ayant une connaissance plus détaillée de l'anatomie que Albertosaurus est disponible pour la plupart des autres tyrannosauridés. La découverte de 26 personnes sur un site fournit une preuve de le comportement de meute et permet des études de ontogenèse et la biologie de la population qui sont impossibles avec les dinosaures moins connus.
Description

Albertosaurus était plus petit que d'autres tyrannosaures, comme Tarbosaurus et Tyrannosaurus. Adultes typiques de Albertosaurus mesurés jusqu'à 9 mètres (30 pieds) de long, tandis que les personnes de grand âge rares pourraient atteindre plus de 10 mètres (33 pi) de longueur. Plusieurs estimations de masse indépendants, obtenus par des méthodes différentes, suggèrent qu'un adulte pèse entre 1,3 Albertosaurus (1,4 tonnes tonnes courtes) et 1,7 tonnes (1,9 tonnes).
Tous, y compris tyrannosauridés Albertosaurus, partageaient une apparence physique similaire. Typiquement pour un théropode, Albertosaurus était bipède et équilibrée la tête lourde et torse avec une longue queue. Cependant, les membres antérieurs de tyrannosauridés étaient extrêmement petite pour la taille de leur corps et conservés que deux chiffres. Les membres postérieurs étaient longues et terminé à un pied à quatre doigts. Le premier chiffre, appelé le hallux, était court et que les trois autres ont touché le sol, avec le troisième (milieu) chiffres plus que le reste. Albertosaurus peut-être été en mesure d'atteindre des vitesses de marche de 14 à 21 kilomètres par heure (8-13 milles à l'heure). Au moins pour les personnes plus jeunes, une grande vitesse de course est plausible.
Crâne et dents

Le massif crâne d'Albertosaurus, perché sur un court, en forme de S cou, était d'environ 1 mètre (3,3 pieds) de long dans les plus grands adultes. De larges ouvertures dans le crâne ( fenestrae) a réduit le poids de la tête tout en fournissant un espace pour fixation musculaire et organes sensoriels. Ses longues mâchoires contenaient, les deux parties combinées, 58 ou plusieurs dents en forme de banane; grandes tyrannosauridés possédaient moins de dents, Gorgosaurus au moins soixante-deux. Contrairement à la plupart des théropodes, Albertosaurus et d'autres tyrannosaures étaient hétérodonte, avec des dents de différentes formes en fonction de leur position dans la bouche. Le dents prémaxillaires à la pointe de la mâchoire supérieure, quatre de chaque côté, étaient beaucoup plus petits que les autres, plus étroitement emballé, et en forme de D dans la coupe transversale. Comme avec Tyrannosaurus, les dents maxillaires (joue) d'Albertosaurus ont été adaptés en forme générale de résister à des forces latérales exercées par la proie du mal. La force de morsure d'Albertosaurus était moins redoutable cependant, la force maximale, par les dents postérieures, atteignant 3,413 Newton. Au-dessus des yeux étaient courtes crêtes osseuses qui peuvent avoir été colorés dans la vie et utilisés dans la cour pour attirer un compagnon.

William Abler observé en 2001 que Albertosaurus dentelures de dents sont si minces pour être fonctionnellement une craquer sous la dent. Cependant, à la base de cette fissure est le vide autour appelé ampoule qui aurait fonctionné à distribuer la force sur une plus grande zone de surface, ce qui entrave la capacité du "crack" formé par la dentelure se propager à travers la dent. L'examen des autres prédateurs anciens, un Phytosaure et Dimetrodon ont également constaté des fissures comme dentelures, mais pas Adaptations pour empêcher la propagation de la fissure. Tyrannosauridés dents ont été utilisés comme des crampons pour tirer la viande hors d'un corps, plutôt que fonctions de découpe couteau semblable. traces d'usure des dents allusion au fait que les comportements de secouer la tête complexes peuvent avoir été impliqué dans l'alimentation tyrannosaure. Quand un tyrannosaure aurait tiré sur un morceau de viande, la force aurait tendance à pousser la pointe de la dent vers l'avant de la bouche et le ancré racine subirait tension sur le côté postérieur et la compression de l'avant. Ce serait typiquement incliner la dent à la formation de fissures sur la côté postérieur de la dent, mais les ampoules à la base des dentelures déjà fissures comme tendraient à diffuser forces d'élite formant potentiels. Cette forme ressemble techniques utilisées par guitare décideurs de «répandre des régions de la flexibilité et de la rigidité d'alterner avec un bâton de bois." L'utilisation d'un foret pour créer une "ampoule" de sortes et de prévenir la propagation de fissures à travers un matériau important est également utilisé pour protéger des surfaces d'avion. Abler a démontré que un bar en plexiglas avec des entailles et des trous forés était plus de 25% plus fort que celui avec incisions ne placées régulièrement.
Classification et de la systématique
| Cladogramme de Tyrannosauridae soulignant la position de Albertosaurus | |||||||||||||||
| |||||||||||||||
Albertosaurus est un membre du théropode Tyrannosauridae famille, dans la sous-famille Albertosaurinae. Son plus proche parent est le un peu plus âgés libratus Gorgosaurus (parfois appelé Albertosaurus libratus; voir ci-dessous). Ces deux espèces sont les albertosaurines que décrites; peuvent exister d'autres espèces non décrites. Thomas Holtz trouvé Appalachiosaurus d'être un albertosaurine en 2004, mais son travail non publié plus récente localise juste à l'extérieur Tyrannosauridae, en accord avec d'autres auteurs.
L'autre sous-famille importante de tyrannosauridés est le Tyrannosaurinae, y compris Daspletosaurus, Tarbosaurus et Tyrannosaurus. Par rapport à ces tyrannosaurines robustes, albertosaurines avaient construit élancée, avec proportionnellement plus petits crânes et os longs de la jambe ( tibia) et les pieds ( métatarses et phalanges).
Découverte et nommage
Appellation
Albertosaurus a été nommé par Henry Fairfield Osborn dans une note d'une page à la fin de 1905 sa description du Tyrannosaurus rex. Les nom honore Alberta, le Province canadienne créée la même année, dans laquelle ont été trouvés les premiers restes. Le nom générique intègre également les grecs terme σαυρος / sauros («lézard»), le suffixe le plus courant dans les noms de dinosaures. Le tapez espèces Albertosaurus est sarcophage; la nom spécifique est dérivé du grec anciennes σαρκοφάγος (sarkophagos) signifie «chair-manger" et ayant la même étymologie comme le récipient funéraire avec lequel il partage son nom: une combinaison des mots grecs σαρξ / sarx («chair») et φαγειν / phagein ("manger"). Plus de trente spécimens de tous les âges sont connus à la science.
Les premières découvertes
Le Type spécimen est un crâne partiel, recueillies à l'été 1884 à partir d'un affleurement du Formation de Horseshoe Canyon à côté de la La rivière Red Deer, en Alberta. Ce spécimen, découvert le 9 Juin 1884, a été récupéré par une expédition de la Commission géologique du Canada, dirigé par le célèbre géologue Joseph Burr Tyrrell. En raison d'un manque d'équipement spécialisé le crâne presque complet pu être que partiellement fixé. En 1889, un collègue de Tyrell Thomas Chesmer Weston a trouvé un crâne plus petit incomplète associée à une certaine matériel squelettique à proximité. Les deux crânes ont été affectés à la incrassatus des espèces préexistantes par Edward Drinker Cope en 1892, bien que le nom était Laelaps préoccupée par un genre de acariens et avait été changé pour Dryptosaurus en 1877 par Othniel Charles Marsh. Cope a refusé de reconnaître le nouveau nom créé par son grand rival Marsh, cependant, Lawrence Lambe lieu de Laelaps incrassatus utilisé le nom Dryptosaurus incrassatus quand il a décrit en détail les restes en 1903 et 1904, une combinaison abord inventé par Oliver Perry Hay en 1902. Peu de temps plus tard, Osborn a souligné que D. incrassatus était basée sur les dents de tyrannosauridés génériques, de sorte que les deux crânes Horseshoe Canyon ne pouvait pas être renvoyé en toute confiance à cette espèce. Les crânes de Horseshoe Canyon différaient également considérablement à partir des restes de D. aquilunguis, espèce type du Dryptosaurus, afin Osborn a créé le nouveau nom Albertosaurus sarcophage pour eux en 1905. Il n'a pas décrit les restes dans le détail, citant description complète de Lambe l'année précédente. Les deux échantillons (le holotype CMN 5600 et la paratype CMN 5601) sont stockés dans la Musée canadien de la nature à Ottawa . Au début du XXIe siècle certaines préoccupations ont surgi qui, en raison de l'état endommagé de l'holotype, Albertosaurus pourrait être un nomen dubium, un «nom douteuse" qui ne peut être utilisé pour le type spécimen se parce que d'autres fossiles pouvaient pas fiable lui attribuer. Cependant, en 2010 Thomas Carr établi que l'holotype, le paratype et comparables trouve tard tous partageaient un seul trait unique ou commune autapomorphie: la possession d'une ouverture agrandie dans le pneumatique arrière de la jante du côté osseuse palatine, prouvant que Albertosaurus était valide taxon.
Dry Island bonebed

Le 11 Août 1910, paléontologue américain Barnum Brown a découvert les restes d'un grand groupe d'Albertosaurus à un autre exploiter en carrière aux côtés de la rivière Red Deer. En raison du grand nombre d'os et le temps limité disponible, le parti de Brown n'a pas recueilli de chaque spécimen, mais fait en sorte de recueillir les restes de tous les individus qu'ils pourraient identifier dans la dépôt d'ossements. Parmi les os déposés dans le Musée américain d'histoire naturelle dans les collections New York City sont sept ensembles de droit métatarsiens, avec deux os des orteils isolés qui ne correspondent pas l'un des métatarses taille. Cela indique la présence d'au moins neuf personnes dans la carrière. Le Royal Tyrrell Museum of Paleontology a redécouvert le dépôt d'ossements en 1997 et a repris le travail de terrain sur le site, qui est maintenant situé à l'intérieur Parc Dry Island Buffalo Jump provincial. Autres fouilles 1997-2005 se les restes de treize individus plus de différents âges, y compris un diminutif de deux ans et une très vieille personne estimée à plus de 10 mètres (33 pi) de longueur. Aucune de ces personnes sont connues de squelettes complets, et la plupart sont représentés par les restes dans les deux musées. Les fouilles ont continué jusqu'en 2008, lorsque le nombre minimum de personnes présentes a été établi à douze ans, sur la base d'éléments conservés qui se produisent qu'une seule fois dans un squelette, et à vingt-six si des éléments ont été comptés en miroir lors de différentes tailles en raison de ontogenèse. Un total de 1128 os Albertosaurus avait été fixé, la plus grande concentration de grands fossiles de théropodes connus du Crétacé.
Gorgosaurus libratus
En 1913, le paléontologue Charles H. Sternberg récupéré une autre tyrannosaurid squelette de l'un peu plus âgés Formation de Dinosaur Park en Alberta. Lawrence Lambe nommé ce dinosaure Gorgosaurus libratus en 1914. D'autres spécimens ont été retrouvés en Alberta et l'État américain du Montana. Trouver, principalement en raison d'un manque de matériel de bonne crâne Albertosaurus, aucune différence significative pour séparer les deux taxons, Dale Russell a déclaré le nom d'un Gorgosaurus synonyme junior de Albertosaurus, qui avait été nommé en premier, et G. libratus a été rebaptisé Albertosaurus libratus en 1970. Une distinction des espèces a été maintenue en raison de la différence d'âge. Cet ajout a élargi la gamme temporelle du genre Albertosaurus arrière par plusieurs millions d'années et son aire de répartition vers le sud par des centaines de kilomètres.
En 2003, Philip J. Currie, bénéficiant de beaucoup plus vastes découvertes et une augmentation générale des connaissances anatomiques des théropodes, comparé plusieurs crânes de tyrannosauridés et est venu à la conclusion que les deux espèces sont plus distincte qu'on ne le pensait. La décision d'utiliser un ou deux genres est plutôt arbitraire, que les deux espèces sont taxons sœur, plus étroitement liés les uns aux autres que de toute autre espèce. Reconnaissant ce fait, Currie a néanmoins recommandé que Albertosaurus et Gorgosaurus être retenus comme des genres séparés, comme il a conclu qu'ils ne étaient plus semblables que Daspletosaurus et Tyrannosaurus, qui sont presque toujours séparés. En outre, plusieurs spécimens albertosaurine ont été récupérés à partir Alaska et Nouveau-Mexique, et Currie ont suggéré que l'Albertosaurus - Situation Gorgosaurus peut être clarifiée une fois ceux-ci sont décrites en détail. La plupart des auteurs ont suivi la recommandation de Currie, mais certains ne ont pas.
Autres découvertes
En 1911, Barnum Brown, au cours de la deuxième année de l'American Museum of Natural History opérations en Alberta, à découvert un crâne Albertosaurus partielle fragmentaire à la rivière Red Deer près de Tolman Bridge, spécimen AMNH 5222.
William Parcs décrit une nouvelle espèce en 1928, Albertosaurus arctunguis, basé sur un squelette partiel manque le crâne fouillé par Gus Lindblad et Ralph Hornell près de la rivière Red Deer en 1923, mais cette espèce a été considérée comme identique à A. sarcophage depuis 1970. spécimen Parcs »(ROM 807) est logé dans le Musée royal de l'Ontario, à Toronto .
Entre 1926 et 1972, aucun fossile Albertosaurus ont été trouvés à tous, mais depuis les années soixante-dix, il ya eu une augmentation constante dans le matériau connu. Outre le dépôt d'ossements Dry Island, six autres crânes et squelettes ont été découverts depuis en Alberta et sont logés dans divers musées canadiens: spécimens RTMP 81.010.001, trouvé en 1978 par le paléontologue amateur Maurice Stefanuk; RTMP 85.098.001, trouvé par Stefanuk le 16 Juin 1985; RTMP 86.64.001 (Décembre 1985); RTMP 86.205.001 (1986); 97.058.0001 RTMP (1996); et la CMN 11315. En raison de vandalisme et accidents cependant, aucune crânes intacts et complets pourrait être assurée entre ces trouvailles. Les fossiles ont également été signalés dans les États américains du Montana, du Nouveau-Mexique, et Wyoming, mais ceux-ci ne représentent probablement pas A. sarcophage et peut même ne pas appartenir au genre Albertosaurus.
Autres espèces
En dehors de A. sarcophage, A. arctunguis et A. libratus, plusieurs autres espèces d'Albertosaurus ont été nommés. Tous ces éléments sont aujourd'hui perçus comme étant plus jeunes synonymes d'autres espèces ou dubia nomina, et ne sont pas affectés à Albertosaurus.
En 1930, Anatoli Nikolaïevitch Riabinin nommé pericolosus Albertosaurus basé sur une dent de la Chine, appartenant probablement à Tarbosaurus. Friedrich von Huene rebaptisé Dryptosaurus incrassatus, pas considéré comme un nomen dubium par lui, à Albertosaurus incrassatus en 1932. Parce qu'il avait identifié Gorgosaurus avec Albertosaurus, Russell en 1970 également renommé Gorgosaurus sternbergi Matthew Brown et 1922 dans un sternbergi Albertosaurus et Gorgosaurus lancensis Gilmore 1946 dans un lancensis Albertosaurus. Les anciens espèce est aujourd'hui considérée comme une forme juvénile de Gorgosaurus libratus, ce dernier comme étant identique à Tyrannosaurus ou représentant un genre distinct Nanotyrannus. Albertosaurus megagracilis a été fondée en 1988 par Gregory S. Paul sur un petit squelette tyrannosaurid, spécimen LACM 28345, de la Formation Hell Creek du Montana. Il a été rebaptisé Dinotyrannus en 1995, mais est maintenant considéré pour représenter un mineur Tyrannosaurus rex. Aussi en 1988, Paul renommé Alectrosaurus olseni Gilmore 1933 dans un Albertosaurus olseni; cela n'a trouvé aucune acceptation générale. En 1989, Gorgosaurus de Maleev 1955 était de Bryn Mader et Robert Bradley renommé dans un novojilovi Albertosaurus; aujourd'hui ce est considéré comme un synonyme de Tarbosaurus.
Sur deux espèces occasions à base de matériau Albertosaurus valables ont été réaffectés à un genre différent: en 1922 William Diller Matthew renommé A. sarcophage Deinodon sarcophage et en 1939, le paléontologue allemand Oskar Kuhn renommé A. arctunguis dans arctunguis Deinodon.
Paléobiologie
modèle de croissance

La plupart des catégories d'âge de Albertosaurus sont représentés dans le registre fossile . Utilisation de l'os histologie, l'âge d'un animal particulier au moment de la mort peut souvent être déterminée, permettant des taux de croissance à estimer et de comparer avec d'autres espèces. Le plus jeune connu Albertosaurus est deux ans découvert dans le dépôt d'ossements Dry Island, qui aurait pesé environ 50 kg (110 lb) mesuré et un peu plus de 2 mètres (7 ft) de longueur. Les 10 mètres (33 pi) spécimen de la même carrière est le plus ancien et le plus grand connu, à 28 ans. Lorsque les spécimens d'âge intermédiaire et la taille sont reportés sur un graphique, un S en forme de résultats de courbe de croissance, avec la croissance la plus rapide survenant dans une période de quatre ans se terminant autour de la seizième année de la vie, un modèle aussi vu dans d'autres tyrannosaures. Le taux de croissance pendant cette phase était de 122 kilogrammes (£ 268) par an, sur la base d'un adulte 1.3 (1,4 tonnes tonnes courtes). D'autres études ont suggéré un poids plus élevé adultes; cela affecterait la grandeur de la vitesse de croissance, mais pas la structure globale. Tyrannosauridés de taille similaire à Albertosaurus avaient des taux de croissance similaires, bien que le Tyrannosaurus rex beaucoup plus a augmenté presque cinq fois plus rapides (601 kg [£ 1325] par an) à son apogée. La fin de la phase de croissance rapide suggère l'apparition de la maturité sexuelle dans Albertosaurus, bien que la croissance se est poursuivie à un rythme plus lent tout au long de la vie des animaux. Maturation sexuelle tout en pleine croissance semble être un trait partagé entre petits et grands dinosaures ainsi que dans les grands mammifères tels que les humains et les éléphants . Ce modèle de la maturation sexuelle relativement tôt diffère frappante du modèle chez les oiseaux, qui retardent leur maturité sexuelle avant d'avoir fini de grandir.
Pendant la croissance, grâce à l'épaississement de la morphologie de la dent tellement changé que, si l'association de squelettes jeunes et adultes sur le dépôt d'ossements Dry Island pas prouvé qu'ils appartenaient à la même taxon, les dents des mineurs auraient probablement été identifié par l'analyse statistique que ceux de une espèce différente.
Histoire de vie
La plupart des personnes ont été connus Albertosaurus âgés de 14 ans ou plus au moment de la mort. Animaux juvéniles sont rarement trouvés sous forme de fossiles, pour plusieurs raisons, principalement biais de la préservation, où les petits os des animaux plus jeunes étaient moins susceptibles d'être préservés par la fossilisation que les grandes os des adultes, et le biais de collecte, où les fossiles plus petits sont moins susceptibles d'être remarqué par les collectionneurs dans le domaine. Jeune Albertosaurus sont relativement grande pour les jeunes animaux, mais leurs restes sont encore rares dans les archives fossiles par rapport aux adultes. Il a été suggéré que ce phénomène est une conséquence de l'histoire de la vie, plutôt que de biais, et que les fossiles des juvéniles Albertosaurus sont rares tout simplement parce qu'ils ne sont pas morts aussi souvent que les adultes ont fait.
Une hypothèse de l'histoire de la vie Albertosaurus postule que nouveau-nés sont morts en grand nombre, mais ne ont pas été conservés dans les archives fossiles en raison de leur petite taille et de la construction fragile. Après seulement deux ans, les mineurs étaient plus grandes que tout autre prédateur dans la région en dehors de Albertosaurus adultes, et plus agile que la plupart de leurs proies. Cela a entraîné une baisse spectaculaire de leur taux de mortalité et une rareté correspondant de restes fossiles. Les taux de mortalité a doublé à douze ans, peut-être le résultat des exigences physiologiques de la phase de croissance rapide, puis de nouveau doublé avec le début de la maturité sexuelle entre l'âge de quatorze à seize ans. Ce taux élevé de mortalité a continué à l'âge adulte, peut-être en raison des exigences physiologiques élevés de la procréation, y compris le stress et les blessures reçues au cours de la compétition intraspécifique pour les compagnons et les ressources, et finalement, les effets toujours croissants de sénescence. Le taux de mortalité plus élevé chez les adultes peut expliquer leur conservation plus fréquente. Très grands animaux étaient rares parce que peu de personnes ont survécu assez longtemps pour atteindre ces tailles. Les taux de mortalité infantile élevés, suivis par réduction de la mortalité chez les jeunes et une augmentation soudaine de la mortalité après la maturité sexuelle, avec très peu d'animaux qui atteignent la taille maximale, est une tendance observée dans de nombreux grands mammifères modernes, y compris des éléphants, des buffles d'Afrique , et rhinocéros. La même tendance est également observée dans d'autres tyrannosaures. La comparaison avec les animaux modernes et d'autres tyrannosauridés apporte son soutien à cette histoire hypothèse de la vie, mais un biais dans les archives fossiles peut encore jouer un rôle important, d'autant plus que plus des deux tiers de tous les spécimens Albertosaurus sont connus d'une localité.
comportement Pack
Le dépôt d'ossements Dry Island découvert par Barnum Brown et son équipage contient les restes de 26 Albertosaurus, les la plupart des individus trouvés dans une localité de ne importe quel grand théropode du Crétacé, et la deuxième plus grande de toute dinosaure théropode derrière l' Allosaurus assemblage au Cleveland Lloyd Dinosaur Quarry Utah. Le groupe semble être composé d'un très vieux adultes; huit adultes entre 17 et 23 ans; sept sous-adultes en leurs phases de croissance rapide à entre 12 et 16 ans; et six mineurs entre les âges de 2 et 11 ans, qui ne avaient pas encore atteint la phase de croissance.

La quasi-absence d' herbivores dépouille et le même état de conservation commune aux nombreuses personnes au dépôt d'ossements carrière Albertosaurus conduit Currie à conclure que la localité ne était pas un piège des prédateurs comme le La Brea Tar Pits en Californie , et que tous les animaux sont morts conservés dans le même temps. Currie affirme cela comme une preuve d'un comportement de meute. D'autres scientifiques sont sceptiques, observant que les animaux peuvent avoir été roulé ensemble par la sécheresse, une inondation ou pour d'autres raisons.
Il existe de nombreuses preuves d'un comportement grégaire chez les dinosaures herbivores, y compris cératopsiens et hadrosaures. Toutefois, ne sont que rarement tant de prédateurs de dinosaures trouvés sur le même site. Petits théropodes comme Deinonychus , Coelophysis et Megapnosaurus (Syntarsus rhodesiensis) ont été trouvés dans les agrégations, de même que les grands prédateurs comme Allosaurus et Mapusaurus. Il existe certaines preuves d'un comportement grégaire dans d'autres tyrannosauridés ainsi. Restes fragmentaires de petits individus ont été trouvés aux côtés de "Sue", le Tyrannosaurus monté dans le Field Museum d'Histoire Naturelle de Chicago , et un dépôt d'ossements dans le Formation de Two Medicine du Montana contient au moins trois spécimens de Daspletosaurus, conservés aux côtés de plusieurs hadrosaures. Ces résultats peuvent corroborer la preuve d'un comportement social dans Albertosaurus, bien que certains ou la totalité des localités ci-dessus peuvent représenter agrégations temporaires ou non naturels. D'autres ont émis l'hypothèse que, au lieu des groupes sociaux, au moins certaines de ces découvertes représentent Dragon de Komodo comme mobbing des carcasses, où la concurrence agressive conduit à certains des prédateurs étant tué et cannibalisé.
Currie offre également la spéculation sur les habitudes pack-chasse de Albertosaurus. Les proportions branche des individus plus petits étaient comparables à ceux de ornithomimidés, qui étaient probablement parmi les plus rapides des dinosaures. Jeune Albertosaurus était probablement tout aussi agiles, ou du moins plus vite que leur proie. Currie émis l'hypothèse que les plus jeunes membres de la meute peuvent avoir été responsable de la conduite de leurs proies vers les adultes, qui étaient plus gros et plus puissant, mais aussi plus lent. Les mineurs peuvent aussi avoir eu différents modes de vie que les adultes, remplissage prédateur niches entre les adultes et les énormes théropodes contemporains plus petits, le plus grand des deux qui étaient ordres de grandeur plus petite que la masse des adultes Albertosaurus. Une situation similaire est observée dans les dragons de Komodo modernes, avec des nouveau-nés à compter vie comme petit insectivores avant de croître pour devenir les prédateurs dominants sur leurs îles. Cependant, comme la préservation de comportement dans les archives fossiles est extrêmement rare, ces idées ne peuvent pas facilement être testés. En 2010, Currie, bien que toujours en faveur de l'hypothèse de meute, a admis que la concentration aurait pu être provoquée par d'autres causes, telles que le niveau d'eau monte lentement au cours d'une inondation prolongée.
Paléopathologie
En 2009, les chercheurs ont émis l'hypothèse que les trous lisses tranchants trouvés dans les mâchoires fossiles de dinosaures tyrannosauridés comme Albertosaurus ont été causés par un parasite semblable à Trichomonas gallinae qui infecte les oiseaux. Ils ont suggéré que tyrannosauridés transmis l'infection en piquant l'autre, et que l'infection altérées leur capacité à manger de la nourriture.
En 2001, Bruce Rothschild et d'autres ont publié une étude examinant la preuve pour fractures de stress et avulsions du tendon dans théropodes dinosaures et les implications pour leur comportement. Ils ont constaté qu'un seul des 319 os Albertosaurus pieds vérifiés pour les fractures de stress entre eux avaient fait et aucun des quatre os de la main fait. Les scientifiques ont constaté que les fractures de stress étaient "significativement" moins fréquente dans Albertosaurus que dans le carnosaure Allosaurus . ROM 807, l'holotype de A. arctunguis (maintenant appelé A. sarcophage), avaient un trou profond de 2,5 par 3,5 cm dans le lame iliaque, bien que le descripteur de l'espèce ne reconnaît pas cela comme pathologique. L'échantillon contient également certaines exostose au quatrième métatarsien gauche. Deux des cinq spécimens Albertosaurus sarcophage avec humérus en 1970 ont été signalés par Dale Russel comme ayant lésions pathologiques pour eux.
En 2010, la santé de l'ensemble Dry Island Albertosaurus a été signalé sur. La plupart des spécimens ont montré aucun signe de maladie. Sur trois phalanges du pied étrange éperons osseux, comprenant des ossification anormale des tendons, dite enthésophytes, étaient présents, leur cause inconnue. Deux côtes et le ventre-côtes ont montré des signes de rupture et de la guérison. Un spécimen adulte avaient une mâchoire inférieure gauche montrant une plaie perforante et les deux marques de morsure guéries et non cicatrisées. Le faible nombre d'anomalies se compare favorablement à l'état de santé d'un Majungasaurus population dont il a été établi en 2007 que 19% des individus a montré pathologies osseuses.
Paléoécologie

Tous les fossiles identifiables de Albertosaurus sarcophagus sont connus de la partie supérieure Formation de Horseshoe Canyon en Alberta. Ces unités plus jeunes de cette géologique date de la formation du début Maastrichtien stade de la Crétacé supérieur, 70-68 Ma (millions d'années). Immédiatement au-dessous de cette formation est le Bearpaw Shale, un formation marin représentant une partie de la Voie maritime intérieure de l'Ouest. La voie maritime reculait que le climat se est refroidi et les niveaux de la mer calmée vers la fin du Crétacé, exposant terres qui avaient été sous l'eau. Mais il ne était pas un bon déroulement du processus, et la voie maritime serait périodiquement élever à des parties de couverture de la région tout au long de Horseshoe Canyon fois avant de finalement recul tout à fait dans les années après. En raison des niveaux de la mer évolution, de nombreux environnements différents sont représentés dans la formation de Horseshoe Canyon, y compris les habitats maritimes à l'offshore et à proximité des côtes et des habitats côtiers comme lagunes, estuaires et battures. De nombreuses charbon représentent coutures ancienne tourbe marécages. Comme la plupart des autres vertébrés fossiles de la formation, Albertosaurus restes sont trouvés dans des dépôts prévues par la deltas et plaines inondables des grands fleuves au cours de la deuxième moitié de Horseshoe Canyon fois.
Le la faune de la formation de Horseshoe Canyon est bien connu, comme des fossiles de vertébrés, y compris ceux des dinosaures, sont très fréquentes. Sharks , rayons, esturgeons, poisson-castor, gars et de la gar-like Aspidorhynchus constitué la faune piscicole. Mammifères inclus multituberculés et marsupial Didelphodon. L'eau salée plésiosaure Leurospondylus a été trouvé dans les sédiments marins dans le Horseshoe Canyon, tandis que les environnements d'eau douce ont été peuplées par des tortues , Champsosaurus, et crocodiliens comme Leidyosuchus et Stangerochampsa. Dinosaures dominent la faune, en particulier les hadrosaures, qui représentent la moitié de tous les dinosaures connus, y compris les genres Edmontosaurus, Saurolophus et Hypacrosaurus. Cératopsiens et ornithomimidés étaient aussi très commune, l'ensemble constitue un autre tiers de la faune connus. Avec beaucoup plus rare ankylosaurians et pachycéphalosaure, tous ces animaux aurait été la proie d'un large éventail de théropodes carnivores, y compris troodontidés, dromaeosaurids, et caenagnathids. Mêlés à la Albertosaurus reste du dépôt d'ossements Dry Island, les os du petit théropode Albertonykus ont été trouvés. Adulte Albertosaurus était le prédateurs dans cet environnement, avec des niches intermédiaires éventuellement remplies par albertosaurs mineurs.