
Dimetrodon
À propos de ce écoles sélection Wikipedia
SOS croit que l'éducation donne une meilleure chance dans la vie des enfants dans le monde en développement aussi. SOS Enfants a regardé des enfants en Afrique depuis quarante ans. Pouvez-vous aider dans leur travail en Afrique ?
| Dimetrodon Plage temporelle: Permien , 299-270Ma | |
|---|---|
 | |
| Squelette monté de Dimetrodon grandis | |
| Classification scientifique | |
| Uni: | Animalia |
| Embranchement: | Chordata |
| Famille: | † Sphenacodontidae |
| Sous-famille: | † Sphenacodontinae |
| Genre: | † Dimetrodon Cope, 1878 |
| Espèce-type | |
| † Dimetrodon limbatus (Cope, 1877 [limbatus origine Clepsydrops]) | |
| Espèce | |
Voir ci-dessous | |
| Synonymes | |
Bathyglyptus Case, 1911 | |
Dimetrodon ( / ré aɪ m ɛ t r ə ré ɒ n /; signifie "deux mesures de dents») est une éteinte genre de synapsid qui a vécu pendant le début du Permien , il ya environ 299 à 270.000.000 années (Ma). Ce est un membre de la famille Sphenacodontidae. La caractéristique la plus marquante de Dimetrodon est la grande voile sur son dos formé par des épines allongées se étendant des vertèbres. Il marchait à quatre pattes et avait un crâne haut, incurvé avec de grandes dents de différentes tailles placés le long des mâchoires. La plupart des fossiles ont été trouvés dans le sud-ouest des États-Unis, la majorité provenant d'un dépôt géologique appelé Lits rouges au Texas et en Oklahoma. Plus récemment, les fossiles ont été trouvés en Allemagne. Plus d'une douzaine d'espèces ont été nommés depuis le genre a été le premier décrit en 1878.
Dimetrodon est souvent confondu avec un dinosaure ou un contemporain des dinosaures dans la culture populaire, mais il a disparu à environ 40 millions d'années avant l'apparition de la première dinosaure dans le Trias période. Généralement reptile comme en apparence et la physiologie, Dimetrodon est néanmoins plus étroitement liée aux mammifères que de ne importe quel groupe de reptiles vivant, si ce ne est pas un ancêtre direct de tous les mammifères. Dimetrodon appartient à un groupe traditionnellement appelé " mammifères comme les reptiles ", plus récemment appelées" tige-mammifères »ou« non mammifère synapsides "parce que beaucoup paléontologues vertébrés groupe aujourd'hui Dimetrodon avec mammifères dans un groupe ou évolutive clade appelé Synapsida tout dinosaures vont de pair avec les reptiles et les oiseaux vivant dans un groupe distinct, Sauropsida. Une grande ouverture unique de chaque côté de l'arrière du crâne Dimétrodon relie les mammifères et la distingue de la plupart des premiers sauropsids, qui soit ouvertures ont peu ou pas deux ouvertures. Des fonctions telles que des nervures à l'intérieur de la la cavité nasale et une crête à l'arrière de la mâchoire inférieure sont considérés comme partie d'une progression évolutive à partir de début tétrapodes (vertébrés à quatre ébranchés) pour les mammifères.
Dimetrodon était probablement l'un des plus grands prédateurs dans les écosystèmes au début du Permien, se nourrissant de poissons et tétrapodes, notamment des reptiles ainsi que les amphibiens. Les petites espèces peuvent Dimetrodon ont eu différents rôles écologiques. La voile de Dimetrodon peut avoir été utilisé pour stabiliser sa colonne vertébrale ou pour chauffer et refroidir son corps comme une forme de thermorégulation. Certaines études récentes affirment que la voile aurait été inefficace à éliminer la chaleur du corps, et a été utilisé dans plus de chances affichage sexuelle.
Description et paléobiologie
Dimetrodon était un synapsid de voile soutenu quadrupède. La plupart des espèces Dimetrodon variaient de longueur de 1,7 à 3,2 mètres (5,6 à 10 pi) et sont estimés avoir pesé entre 28 et 250 kg (62 et £ 550). La plus grande espèce connue de Dimetrodon est D. grandis à 3,2 mètres (10 pi) et le plus petit est D. teutonis à 60 centimètres (24 po). Les plus grandes espèces de Dimetrodon ont été parmi les plus grands prédateurs de la Permien inférieur, bien que la étroitement liés Tappenosaurus, connu à partir de fragments du squelette légèrement roches plus jeunes, peut-être encore plus avec une estimation de 18 pieds (5,5 m) de longueur totale du corps.
Crâne

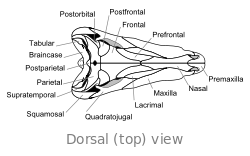
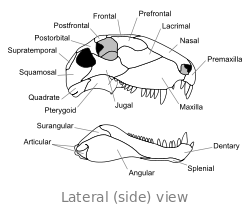
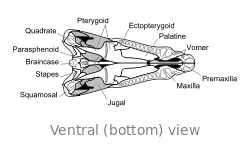
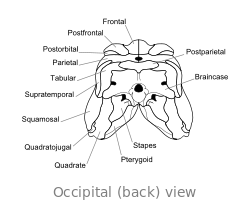
Le crâne de Dimetrodon est grand et comprimé latéralement, ou côte à côte. Les orbites sont positionnés de haut et loin dans le crâne. Derrière chaque orbite de l'œil est un seul trou appelé Fenestra infratemporale. Deux paires de trous supplémentaires peuvent être vus à l'arrière du crâne en vue de dessus: la Fenestra supratemporal et Fenestra posttemporal. L'arrière du crâne (la occiput) est orientée à un léger angle vers le haut, une caractéristique qu'elle partage avec tous les autres synapsides début. Le bord supérieur de la pente vers le bas dans le crâne d'un arc convexe vers l'extrémité du museau. L'extrémité de la mâchoire supérieure, formée par la prémaxillaire os, est soulevée au-dessus de la partie de la mâchoire formée par la maxillaire osseuse pour former une "étape." maxillaire Dans cette étape est un diastème ou lacune dans la rangée de dents.
Dents
La taille de dents diffère grandement sur la longueur des mâchoires, prêt Dimetrodon son nom, qui signifie «deux formes de dents" en référence à des ensembles de petites et de grandes dents. Une ou deux paires de caniniforms (large pointu canines-comme) se étendent depuis le maxillaire. Grandes incisives sont également présents à l'extrémité des mâchoires supérieure et inférieure, enracinée dans la prémaxillaires et os du dentaire. Petites dents sont présents autour de la «étape» maxillaire et derrière les caniniforms, devenant plus petit plus loin dans la mâchoire.
Beaucoup de dents sont plus large à leurs midsections et étroit proche des mâchoires, leur donnant l'apparence d'une larme. Dents en forme de larme sont uniques à dimetrodon et d'autres étroitement liés sphenacodontids, et les aider à distinguer des autres synapsides début. Comme dans de nombreux autres synapsides début, les dents de la plupart des espèces Dimetrodon sont dentelées sur leurs bords. Les dents de D. teutonis manquer dentelures, mais qui ont encore des arêtes vives.
Cavité nasale
Sur la surface intérieure de la partie nasale du crâne sont appelés arêtes nasoturbinals, qui peuvent avoir pris en charge cartilage que l'augmentation de la superficie de la épithélium olfactif, la couche de tissu qui détecte les odeurs. Ces crêtes sont beaucoup plus petits que ceux de synapsides plus tard de la fin du Permien et du Trias, dont la grande nasoturbinals sont pris comme une preuve de sang chaud, car ils peuvent ont soutenu muqueuses qui réchauffait et humidifié l'air entrant. Ainsi, la cavité nasale est de Dimétrodon transition entre ceux des vertébrés et de mammifères terrestres début.
Jaw conjointe et l'oreille
Une autre caractéristique de transition de Dimetrodon est une crête à l'arrière de la mâchoire appelé la lame réfléchie. La lame se trouve réfléchi sur la os articulaire, qui se connecte à la carré os du crâne, pour former l'articulation de la mâchoire. Dans ancêtres mammifères plus tard, le carré articulaire et séparée de l'articulation de la mâchoire pendant que le articulaire développées dans le marteau os de l'oreille interne. La lame réfléchie est devenu partie d'un noyau appelé l'anneau du tympan qui soutient le tympan chez tous les mammifères vivants.
Queue

La queue de Dimetrodon constitue une grande partie de sa longueur totale du corps et comprend environ 50 vertèbres caudales. Tails ont été absents ou incomplets dans les squelettes première décrit des Dimetrodon; la seule vertèbres caudales connu était le onze plus proche de la hanche. Depuis ces quelques premières vertèbres caudales étroite rapidement à mesure qu'ils progressent plus loin de la hanche, de nombreux paléontologues à la fin du XIXe et début du XXe siècles pensé que Dimetrodon avait une queue très courte. Ce ne est qu'en 1927 que la queue largement complète de Dimetrodon a été décrit.
Voile
La voile de Dimetrodon est formé par allongée neurépines saillie à partir de la vertèbre. Chaque colonne varie en forme de section transversale de sa base jusqu'à sa pointe dans ce qui est connu comme la différenciation "dimetrodont". Près du corps de la vertèbre, la section de la colonne vertébrale croix est comprimé latéralement dans une forme rectangulaire, et plus proche de la pointe, il prend une forme en huit comme une rainure longe chaque côté de la colonne vertébrale. La figure-huit forme est pensé pour renforcer la colonne vertébrale, de flexion et de prévenir les fractures. Une section de la colonne vertébrale d'un spécimen de giganhomogenes Dimetrodon est de forme rectangulaire, mais conserve en huit anneaux en forme à proximité de son centre, ce qui indique que la forme des épines peut changer à mesure que les personnes vieillissent. L'anatomie microscopique de chaque pointe varie de la base au sommet, indiquant l'endroit où il a été incorporé dans les muscles du dos et où il a été exposé dans le cadre d'une voile. La partie inférieure ou partie proximale de la colonne vertébrale présente une surface rugueuse qui ont servi de point d'ancrage pour le épaxiale muscles du dos, et dispose également d'un réseau de tissus conjonctifs appelés Les fibres de Sharpey qui indiquent qu'il a été incorporé dans le corps. Plus haut sur la distale (extérieure) de la partie colonne vertébrale, la surface de l'os est plus lisse. Le périoste, une couche de tissu entourant l'os, est couvert dans les petites rainures qui supportaient probablement les vaisseaux sanguins qui vascularisé la voile. La grande rainure qui se étend le long de la colonne vertébrale a été autrefois considéré comme un canal pour les vaisseaux sanguins, mais depuis l'os ne contient pas de canaux vasculaires, la voile ne est pas pensé pour avoir été comme hautement vascularisé que l'on pensait. Certains spécimens de Dimetrodon préserver les zones déformées des épines neurales qui semblent être cicatrisée fractures. Le os cortical qui a grandi au cours de ces pauses est très vascularisé, ce qui suggère que des tissus mous doit avoir été présent sur la voile pour alimenter le site avec des vaisseaux sanguins. Layered os lamellaire constitue l'essentiel de la zone de section transversale de la colonne vertébrale de neurones, et contient des lignes de croissance arrêtés qui peuvent être utilisées pour déterminer l'âge de chaque individu à la mort. Dans de nombreux spécimens les parties distales des épines plier le, indiquant que la voile aurait eu un profil irrégulier dans la vie. Leur malhonnêteté suggère que des tissus mous peut ne pas avoir étendu tout le chemin à l'extrémité des épines, ce qui signifie que la sangle de la voile peut-être pas été aussi étendue comme il est communément imaginé.
Fonction thermorégulatrice

Les paléontologues ont proposé de nombreuses façons dont la voile aurait pu fonctionner dans la vie. Certains des premiers à penser à son but suggéré que la voile peut avoir servi de camouflage parmi les roseaux tout Dimetrodon attendait une proie ou comme une voile de bateau comme réelle pour attraper le vent alors que l'animal était dans l'eau. Une autre est que les longues neurépines pourraient se sont stabilisés en limitant le coffre vers le haut et vers le bas le mouvement, ce qui permettrait un mouvement plus efficace côté à l'autre pendant la marche. En 1940, Alfred Romer et Llewellyn Ivor Prix proposé que la voile a servi une fonction thermorégulatrice, permettant aux individus de se réchauffer leurs corps avec la chaleur du soleil. Dans les années suivantes, de nombreux modèles ont été créés pour estimer l'efficacité de la thermorégulation dans Dimetrodon. Par exemple, dans un article de 1973 dans la revue Nature, les paléontologues CD Bramwell et PB Fellgett estime qu'il a fallu 200 kg (440 lb) individuels sur les heures et demi pour sa température de corps se élever de 26 à 32 ° C (79 à 90 ° F). En 1986, Steven C. Haack a conclu que le réchauffement a été plus lente qu'on ne le pensait et que le processus a probablement fallu quatre heures. En utilisant un modèle basé sur une variété de facteurs environnementaux et les aspects physiologiques hypothétiques de Dimetrodon, Haack a constaté que la voile a permis Dimetrodon se réchauffer plus vite le matin et atteindre une température du corps légèrement plus élevé pendant la journée, mais qu'il était inefficace pour libérer l'excès de chaleur et ne pas laisser Dimetrodon de conserver une température corporelle plus élevée la nuit. En 1999, un groupe d'ingénieurs mécaniques a créé un modèle informatique pour analyser la capacité de la voile à réguler la température du corps pendant les différentes saisons, et a conclu que la voile a été bénéfique pour la capture et libérant de la chaleur à tout moment dans l'année.
La plupart de ces études donnent deux rôles de thermorégulation pour la voile de Dimetrodon: une comme un moyen de réchauffer rapidement le matin, et l'autre comme un moyen pour refroidir lorsque la température du corps devient élevée Dimetrodon et tous les autres premiers vertébrés terrestres Permien sont supposés. ont été de sang-froid ou poïkilothermes, en se appuyant sur le soleil pour maintenir une température corporelle élevée. En raison de sa grande taille, Dimetrodon avait haute inertie thermique, ce qui signifie que les variations de la température du corps ont été plus lentement en ce que chez les animaux plus petits de corps. Comme les températures ont augmenté dans les matins, le petit corps proie des Dimetrodon pourrait se réchauffer leurs corps beaucoup plus rapidement que quelque chose pouvait la taille de Dimetrodon. De nombreux paléontologues y compris Haack ont proposé que la voile de Dimetrodon peut-être permis de se réchauffer rapidement le matin pour suivre le rythme avec sa proie. Grande surface de la voile signifiait aussi la chaleur pourrait se dissiper rapidement dans l'environnement, utile si l'animal nécessaire pour libérer l'excès de chaleur qui a été produite par le métabolisme ou qui a été absorbée par le soleil. Dimetrodon peut avoir un angle sa voile loin du soleil pour se rafraîchir ou restreint le flux sanguin vers le voile de conserver la chaleur pendant la nuit.
En 1986, J. Scott Turner et C. Richard Tracy proposé que l'évolution d'une voile en Dimetrodon était liée à l'évolution de sang chaud chez les ancêtres de mammifères. Ils pensaient que la voile de Dimetrodon lui a permis d'être homéotherme, en maintenant une constante, bien que faible, la température du corps. Les mammifères sont également homéothermes, même si elles diffèrent de Dimetrodon d'être endothermique, contrôler leur température corporelle interne par le métabolisme accru. Turner et Tracy ont noté que les longues jambes de début thérapsides, un groupe plus avancé de synapsides qui sont étroitement liés à des mammifères, leur a permis de libérer la chaleur d'une manière similaire à la voile de Dimetrodon. Le homéothermie qui se est développé chez les animaux comme Dimetrodon peut-être reporté à thérapsides travers une modification de la forme du corps, qui finissent par développer dans le sang chaud des mammifères.
Des études récentes sur la voile de Dimetrodon et d'autres soutiennent sphenacodontids 1986 affirmation de Haack que la voile était mal adapté à dégager de la chaleur et de maintenir une température corporelle stable. La présence de voiles de petites espèces valides de Dimetrodon tels que D. milleri et D. teutonis ne correspond pas à l'idée que le but de la voile était thermorégulation parce que les petites voiles sont moins en mesure de transférer la chaleur et parce que de petits corps peuvent absorber et libérer de la chaleur facilement sur leur propre. En outre, proches parents de Dimetrodon tels que Sphenacodon ont de très faibles crêtes qui aurait été inutile en tant que dispositifs de thermorégulation. La grande voile de Dimetrodon est pensé pour avoir développé progressivement à partir de ces petites crêtes, ce qui signifie que sur la plus grande de l'histoire de l'évolution de la voile, la thermorégulation ne aurait pas servi une fonction importante.
La sélection sexuelle
Grandes spécimens valides de Dimetrodon ont voiles plus grandes par rapport à leur taille, un exemple de allométrie positive. Allométrie positive peut bénéficier thermorégulation car cela signifie que, en tant que personnes deviennent plus grandes, surface augmente plus vite que la masse. Animaux Agrandir valides génèrent une grande quantité de chaleur par le métabolisme, et la quantité de chaleur qui doit être dissipée de la surface du corps est nettement supérieure à ce qui doit être dissipée par des animaux plus petits valides. Dissipation de la chaleur peut être prédit à travers de nombreux animaux différents avec une seule relation entre la masse et la surface. Cependant, une étude de allométrie dans Dimetrodon 2010 a révélé une relation différente entre la voile et la masse corporelle: l'exposant d'échelle réelle de la voile était beaucoup plus grande que l'exposant attendue chez un animal adapté à la dissipation thermique. Les chercheurs ont conclu que la voile de Dimetrodon a augmenté à un rythme beaucoup plus rapide que ce qui était nécessaire pour la thermorégulation, et a suggéré que la sélection sexuelle était la raison principale de son évolution.
Le dimorphisme sexuel


Dimetrodon peut avoir été dimorphisme sexuel, ce qui signifie que les hommes et les femmes avaient des tailles légèrement différentes du corps. Certains spécimens de Dimetrodon ont été émis l'hypothèse que les hommes parce qu'ils ont des os plus épais, de grandes voiles, plus des crânes, et plus prononcés maxillaires «étapes» que d'autres. Sur la base de ces différences, les squelettes montés dans le Musée américain d'histoire naturelle (AMNH 4636) et de la Musée Field d'histoire naturelle peut être des hommes et des squelettes dans le Denver Museum of Nature et Science ( MCZ 1347) et le Université du Michigan Museum of Natural History peut être femelles.
Espèce
| Espèce | Autorité | Emplacement | Statut | Synonymes | Images |
|---|---|---|---|---|---|
| Dimetrodon angelensis | Olson, 1962 | Texas | Valide |  | |
| Dimetrodon booneorum | Romer, 1937 | Texas | Valide | ||
| Dimetrodon cruciger | Cope, 1878 | Texas | Synonyme de Edaphosaurus cruciger | ||
| Dimetrodon dollovianus | Case, 1907 | Texas | Valide | Dollovianus Embolophorus Cope, 1888 | |
| Giganhomogenes Dimetrodon | Case, 1907 | Texas | Valide |  | |
| Gigas Dimetrodon | Cope, 1878 | Texas | Synonyme de Dimetrodon grandis | Clepsydrops gigas Cope, 1878 | |
| Dimetrodon grandis | Romer et Price, 1940 | Oklahoma Texas | Valide | Clepsydrops gigas Cope, 1878 Gigas Dimetrodon Cope, 1878 Theropleura grandis Case, 1907 Cas Bathyglyptus de 1911 Dimetrodon maximus Romer 1936 |  |
| Dimetrodon incisivus | Cope, 1878 | Texas | Synonyme de Dimetrodon limbatus | ||
| ? Dimetrodon kempae | Romer, 1937 | Texas | Possible nomen dubium | ||
| Dimetrodon limbatus | Romer et Price, 1940 | Oklahoma Texas | Valide | Clepsydrops limbatus Cope, 1877 Dimetrodon incisivus Cope, 1878 Dimetrodon rectiformis Cope, 1878 Dimetrodon semiradicatus Cope, 1881 | |
| Dimetrodon longiramus | Case, 1907 | Texas | Synonyme de Obtusidens Secodontosaurus | ||
| Dimetrodon loomisi | Romer, 1937 | Texas Oklahoma | Valide |  | |
| Dimetrodon macrospondylus | Case, 1907 | Texas | Valide | Clepsydrops macrospondylus Cope, 1884 Dimetrodon platycentrus Case, 1907 | |
| Dimetrodon milleri | Romer, 1937 | Texas | Valide |  | |
| Dimetrodon natalis | Romer, 1936 | Texas | Valide | Clepsydrops Cope Natalis, 1878 |  |
| Dimetrodon occidentalis | Berman, 1977 | Arizona Nouveau Mexique Utah | Valide | ||
| Dimetrodon platycentrus | Case, 1907 | Texas | Synonyme de Dimetrodon macrospondylus | ||
| Rectiformis Dimetrodon | Cope, 1878 | Texas | Synonyme de Dimetrodon limbatus | ||
| Dimetrodon semiradicatus | Cope, 1881 | Texas | Synonyme de Dimetrodon limbatus | ||
| Teutonis Dimetrodon | Berman et al., 2001 | Allemagne | Valide |
l'histoire de classification
Premières descriptions par Cope
Fossiles maintenant attribués à Dimetrodon ont été étudiés par le paléontologue américain Edward Drinker Cope dans les années 1870. Cope avait obtenu les fossiles avec ceux de beaucoup d'autres tétrapodes (vertébrés Permien-quatre ébranchés) de plusieurs collectionneurs qui avaient été explorent un groupe de rochers au Texas appelé Lits Rouge. Parmi ces collectionneurs étaient naturaliste suisse Jacob Boll, Texas géologue WF Cummins et paléontologue amateur Charles Hazelius Sternberg. La plupart des spécimens de Cope allés au Musée américain d'histoire naturelle ou à la Université de Walker Museum de Chicago (la plupart de la collection de fossiles Walker est maintenant logé dans le Musée Field d'histoire naturelle).
Sternberg a envoyé certains de ses propres échantillons à paléontologue allemand Ferdinand Broili au Université de Munich, bien que Broili ne était pas aussi prolifique que Cope quand il est venu à décrire spécimens. Cope rival Othniel Charles Marsh a également recueilli quelques os de Dimetrodon, qu'il envoya au Musée Walker. La première utilisation du nom Dimetrodon venu en 1878 lorsque Cope nommé espèces Dimetrodon incisivus, rectiformis Dimetrodon et gigas Dimetrodon dans la revue scientifique Actes de l'American Philosophical Society.
Cependant, la première description d'un fossile Dimetrodon est venu un an plus tôt, lorsque Cope nommé les espèces limbatus Clepsydrops des lits rouges du Texas. (Le nom a été inventé Clepsydrops par Cope en 1875 pour sphenacodontid reste de Vermilion County, Illinois, et a ensuite été utilisé pour de nombreux spécimens sphenacontid du Texas; de nombreuses nouvelles espèces de sphenacodontids du Texas ont été affectés soit Clepsydrops ou Dimetrodon la fin du XIXe et début du XXe siècles.) C. limbatus a été reclassé comme une espèce de Dimetrodon en 1940, ce qui signifie que 1 877 papier Cope était le premier enregistrement de Dimetrodon.
Cope fut le premier à décrire un synapsid de voile soutenu avec la désignation du Clepsydrops natalis dans son article 1878, même se il a appelé la voile un aileron et il par rapport aux crêtes de la modernité lézard basilic (Basilicus). Sails ne ont pas été conservés dans les spécimens de D. incisivus et D. gigas que Cope décrits dans son document de 1878, mais épines allongées étaient présents dans le D. rectiformis spécimen qu'il a décrit. Cope a commenté sur le but de la voile en 1886, écrit, "L'utilitaire est difficile à imaginer. Sauf l'animal avait des habitudes aquatiques, et a nagé sur le dos, la crête ou dérive doivent avoir été dans la façon de mouvements actifs ... Les membres ne sont pas assez longtemps, ni les griffes suffisamment aigus pour démontrer habitudes arboricoles, comme dans le genre existante Basilicus, où une crête similaire existe ".
Les premières descriptions du 20e siècle
Dans les premières décennies du XXe siècle, paléontologue américain Case CE auteur de nombreuses études sur Dimetrodon et décrit plusieurs nouvelles espèces. Il a reçu un financement de la Institution Carnegie pour son étude d'un grand nombre Dimetrodon spécimens dans les collections de la Musée américain d'histoire naturelle et de plusieurs autres musées. Beaucoup de ces fossiles avaient été recueillis par Cope, mais ne avaient pas été bien décrit, que Cope était connu pour ériger de nouvelles espèces sur la base de seulement quelques fragments d'os.
Depuis la fin des années 1920, le paléontologue Alfred Romer réétudiée nombreux spécimens Dimetrodon et nommé plusieurs nouvelles espèces. En 1940, Romer coauteur d'une vaste étude avec Llewellyn Ivor Prix appelé «Examen de la pelycosauria" dans lequel ont été réévalués les espèces de Dimetrodon nommés par Cope et Case. La plupart des noms d'espèces jugé valable par Romer et le prix sont encore utilisés aujourd'hui.
Nouveaux spécimens
Dans les décennies suivantes la monographie de Romer et Prix, de nombreux spécimens Dimetrodon ont été décrits dans des localités autres que le Texas et l'Oklahoma. Le premier a été décrite à partir de la Four Corners région de l'Utah en 1966 et un autre a été décrit de l'Arizona en 1969. En 1975, Olson signalés matériau Dimetrodon de l'Ohio. Une nouvelle espèce de Dimetrodon appelés D. occidentalis (signifiant "ouest Dimetrodon") a été nommé en 1977 au Nouveau-Mexique. Les spécimens trouvés dans l'Utah et de l'Arizona probablement appartiennent également à D. occidentalis.
Avant ces découvertes, une théorie existait qu'une voie maritime du centre du continent séparé ce qui est maintenant le Texas et l'Oklahoma de terres de l'Ouest plus au cours de la début du Permien, isolement Dimetrodon à une petite région d'Amérique du Nord tandis qu'un plus petit appelé sphenacodontid Sphenacodon dominé la zone ouest. Bien que cette voie maritime ne existe sans doute, la découverte de fossiles autres que le Texas et l'Oklahoma montrer que sa mesure était limitée et que ce ne était pas une barrière efficace à la distribution de Dimetrodon.
En 2001, une nouvelle espèce de Dimetrodon appelés D. teutonis a été décrite à partir de la Saar-Nahe bassin de l'Allemagne, l'extension de la portée géographique de Dimetrodon hors Amérique du Nord pour la première fois.
Classification phylogénétique
Dimetrodon est un des premiers membres d'un groupe appelé synapsides, qui comprennent les mammifères et beaucoup de leurs parents disparus. Il est souvent confondu avec un dinosaure dans la culture populaire, en dépit d'avoir disparu à environ 40 millions d'années (Ma) avant la première apparition des dinosaures. En synapsid, Dimetrodon est plus étroitement liée aux mammifères que de dinosaures ou tout reptile vivant. Dans la fin du XIXe et début du XXe siècles, la plupart des paléontologues appellent Dimetrodon un reptile en conformité avec Taxonomie linnéenne. Dans le système de Linné, Reptilia a été classé comme un classe et Dimetrodon a été classé comme un genre au sein de cette classe. Bien qu'une relation étroite entre Dimetrodon et de mammifères a été appréciée à l'époque, les mammifères appartenaient à leur propre catégorie distincte et donc Dimetrodon était un "reptile mammifère-like". Les paléontologues ont reconnu que les mammifères ont évolué à partir de ce groupe dans ce qu'ils appellent une transition reptile à mammifère. Plus récemment, la taxonomie phylogénétique est devenu un système commun de classification en paléontologie des vertébrés où des groupes sont définis par des ancêtres communs partagés. Reptiles et de mammifères ont été placés dans clades, qui sont des groupes qui comprennent un ancêtre évolutive commune et tous ses descendants. Sous la systématique phylogénétique, les descendants de la dernier ancêtre commun des Dimetrodon et tous les reptiles vivant doit inclure tous les mammifères parce Dimetrodon est maintenant connu pour être plus étroitement liée aux mammifères que de tous les reptiles vivant. Pour éviter les mammifères étant dans le clade qui comprend ces reptiles vivant, ni Dimetrodon ni aucune autre "reptile mammifère comme" peuvent être considérés comme faisant partie de ce clade. Descendants du dernier ancêtre commun des mammifères et des reptiles (qui sont apparues autour de 310 Ma dans la Carbonifère tardif) sont donc divisé en deux clades: Synapsida, qui comprend Dimetrodon et les mammifères, et Sauropsida, qui comprend reptiles vivants et tous les reptiles disparus plus étroitement liées à eux que pour les mammifères.
Dans Synapsida, Dimetrodon fait partie du clade Sphenacodontia. Sphenacodontia a d'abord été proposé comme un groupe synapsid au début de 1940 par les paléontologues Alfred Romer et Llewellyn Ivor prix, ainsi que les groupes Ophiacodontia et Edaphosauria. Les trois groupes sont connus de la fin du Carbonifère et Permien inférieur. Romer et Prix les distinguent principalement par postcrâniens fonctionnalités telles que les formes des membres et des vertèbres. Ophiacodontia a été considéré comme le groupe le plus primitif parce que ses membres semblaient le plus reptilienne, et Sphenacodontia était le plus avancé parce que ses membres semblaient le plus comme un groupe appelé Therapsida, qui comprenait les parents les plus proches de mammifères. Romer et Prix placés un autre groupe de début syapsids appelé varanopids sein Sphenacodontia, les considérant comme plus primitif que les autres sphenacodonts comme Dimetrodon. Ils pensaient varanopids et Dimetrodon -comme sphenacodonts ont été étroitement liés parce que les deux groupes étaient carnivores, bien varanopids sont beaucoup plus petits et plus, voiles dépourvus de lézard. La vision moderne des relations synapsid a été proposé par le paléontologue Robert R. Reisz en 1986, dont l'étude Fonctions incluses trouve principalement dans le crâne plutôt que dans le squelette post-crânien. Dimetrodon est toujours considéré comme un sphenacodont en vertu du présent phylogénie, mais varanodontids sont maintenant considérés comme plus synapsides basales, échappant Sphenacodontia. Dans Sphenacodontia est le groupe Sphenacodontoidea, qui à son tour contient Sphenacodontidae et Therapsida. Sphenacodontidae est le groupe contenant Dimetrodon et plusieurs autres synapsides de voile soutenu comme Sphenacodon et Secodontosaurus, tandis que Therapsida comprend les mammifères et leurs essentiellement du Permien et du Trias parents. Voici un cladogramme modifié à partir de l'analyse de Benson (sous presse) qui suit cette phylogénie:
| Amniota |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Paléoécologie

Les fossiles de Dimetrodon sont connus des États-Unis (Texas, Oklahoma, Nouveau-Mexique, l'Arizona, l'Utah et de l'Ohio) et l'Allemagne, les zones qui faisaient partie du supercontinent Euramerica au cours du Permien précoce. Aux États-Unis, presque tout le matériel attribué à Dimetrodon est venue de trois groupes géologiques dans le nord-centre du Texas et le sud-centre de l'Oklahoma: le Clear Fork Groupe, le Wichita Groupe, et de la River Group Pearce. La plupart des découvertes de fossiles font partie des écosystèmes des plaines qui, au cours du Permien, auraient été de vastes zones humides. En particulier, les lits Rouge du Texas est une région d'une grande diversité de fossiles tétrapodes, ou vertébrés à quatre bite. En plus de Dimetrodon, tétrapodes les plus courantes dans les lits rouges et partout premiers dépôts du Permien dans le sud-ouest des États-Unis sont les amphibiens Archeria, Diplocaulus, Eryops, et Trimerorhachis, la reptiliomorph Seymouria, le reptile Captorhinus et les synapsides Ophiacodon et Edaphosaurus. Ces tétrapodes formaient un groupe d'animaux qui paléontologue Everett C. Olson appelé «permo-carbonifère chronofauna,« un la faune qui dominaient l'écosystème euroaméricains continentale pendant plusieurs millions d'années. Basé sur la géologie des dépôts comme les lits rouges, la faune est pensé pour avoir habité une plaine bien végétalisé écosystème deltaïque.
Réseau trophique
Olson a fait beaucoup de déductions sur la paléoécologie des lits Texas Red et le rôle de Dimetrodon au sein de son écosystème. Il a proposé plusieurs principaux types d'écosystèmes dans lesquels les premiers tétrapodes vivaient. Dimetrodon appartenait à l'écosystème le plus primitif, qui se est développée à partir des réseaux trophiques aquatiques. Dans ce document, les plantes aquatiques ont été les producteurs primaires et ont été largement nourri de par les poissons et les invertébrés aquatiques. La plupart des vertébrés terrestres alimentés sur ces consommateurs primaires aquatiques. Dimetrodon était probablement le prédateur de l'écosystème Red Beds, se nourrissant de divers organismes tels que les gros poissons Xenacanthus, les amphibiens aquatiques Trimerorhachis et Diplocaulus, et les tétrapodes terrestres et Seymouria Trematops. Les insectes sont connus depuis le début des lits rouges du Permien et sont probablement impliqués dans une certaine mesure dans la même chaîne alimentaire que Dimetrodon, se nourrissant de petits reptiles comme Captorhinus. L'assemblage Red Beds comprenait également quelques-uns des premiers grands herbivores terrestres vivant comme Edaphosaurus et Diadectes. Nourrir principalement sur les plantes terrestres, ces herbivores ne tirent leur énergie de réseaux trophiques aquatiques. Selon Olson, le meilleur analogue moderne pour la Dimetrodon de l'écosystème habité est le Everglades.
Les seules espèces de Dimetrodon trouvés à l'extérieur du sud-ouest des États-Unis est Dimetrodon teutonis de l'Allemagne. Ses restes ont été trouvés dans le Formation Tambach dans un site fossile appelé la localité Bromacker. L'assemblage de la Bromacker des premiers tétrapodes Permien est inhabituel en ce qu'il existe quelques grandes synapsides valides desservant le rôle des grands prédateurs. D. teutonis est estimée à seulement 1,7 mètres (5,6 pieds) de longueur, trop petit pour proies sur la grande herbivores diadectid qui sont abondants dans l'assemblage Bromacker. Il mangeait plus susceptibles de petits vertébrés et d'insectes. Seuls trois fossiles peuvent être attribués à de grands prédateurs, et ils sont considérés comme ayant été soit de grandes ou de petites varanopids sphenacodonts, deux facteurs qui pourraient potentiellement proies D. teutonis. Contrairement aux plaines deltaïques lits rouges du Texas, les dépôts Bromacker sont considérés comme ayant représenté un environnement de hautes terres sans espèces aquatiques. Il est possible que les grands-bodied carnivores ne faisaient pas partie de l'assemblage Bromacker parce qu'ils étaient dépendantes de grands amphibiens aquatiques pour leur alimentation.
Mineurs
Bien que certaines espèces Dimetrodon pourraient devenir très volumineux, de nombreux spécimens juvéniles sont connus. Paléontologiste Robert Bakker a suggéré en 1982 que les adultes Dimetrodon habitée plaines inondables ouverts tandis que les juvéniles préfère plus marécages et les marges de lac isolé. Ces environnements ont été déduites du type de sédiments dans laquelle ont été trouvés les os Dimetrodon. Bakker pensait que les mineurs Dimetrodon se seraient comportés de manière similaire aux jeunes de vivre de grands reptiles à éviter les adultes de leur propre espèce. Chez les reptiles vivant, les adultes peuvent concourir pour les mêmes ressources que leurs petits et peuvent même leur proie. Cependant, une étude ultérieure par Donald Brinkman jeter le doute sur l'idée de Bakker en montrant que les spécimens trouvés dans les sédiments des plaines inondables et marécageuses représentent deux espèces différentes, pas les populations adultes et juvéniles de la même espèce.