
Eucaryote
À propos de ce écoles sélection Wikipedia
Arrangeant une sélection Wikipedia pour les écoles dans le monde en développement sans internet était une initiative de SOS Enfants. Le parrainage d'enfants aide les enfants un par un http://www.sponsor-a-child.org.uk/ .
| Les cellules eucaryotes Plage temporelle: il ya 1,6 à 2100000000 années (peut-être dès il ya 2,7 milliards d'années) - Présent Had'n Archéen Protérozoïque
Pha. | |
|---|---|
 | |
| Eucaryotes et quelques exemples de leur diversité | |
| Classification scientifique | |
| Domaine: | Eucaryotes Whittaker & Margulis, 1978 |
| Royaumes | |
Animalia - Animaux
Amibes Plantae - Plantes Chromalveolata Rhizaria Excavata | |
| Alternative phylogénie | |
| |
Un eucaryote (pron .: / JU k ær Je . oʊ t / Ewing KARR -ee-OHT ou / JU k ær Je ə t /) Est un organisme dont les cellules contiennent des structures complexes enfermés dans membranes. Eucaryotes peuvent plus officiellement être nommé en tant que taxon Eukarya ou Eucaryotes. La structure liée à la membrane qui fixe définissant des cellules eucaryotes en dehors de des cellules procaryotes est le noyau, ou enveloppe nucléaire, dans lequel le matériel génétique est effectuée. La présence d'un noyau donne eucaryotes leur nom, qui vient du grec ευ (UE, «bon») et κάρυον (Karyon, «noix» ou «noyau»). La plupart des cellules eucaryotes contiennent également d'autres membranaire organites tels que les mitochondries , chloroplastes et la L'appareil de Golgi. Tous les grands organismes complexes sont des eucaryotes, y compris les animaux , les plantes et les champignons . Le groupe comprend également plusieurs organismes unicellulaires.
La division cellulaire chez les eucaryotes est différente de celle dans les organismes sans noyau ( Procaryote). Elle consiste à séparer les dupliqué chromosomes, par des mouvements dirigés par les microtubules. Il existe deux types de processus de division. En mitose, une cellule se divise pour produire deux cellules génétiquement identiques. En méiose, qui est nécessaire dans la reproduction sexuée, une cellule diploïde (ayant deux instances de chaque chromosome, un de chaque parent) subit recombinaison de chaque paire de chromosomes parentaux, puis deux étapes de la division cellulaire, ce qui entraîne quatre cellules haploïdes ( gamètes). Chaque gamète a juste un complément de chromosomes, chacun un mélange unique de la paire correspondante de chromosomes parentaux.
Eucaryotes semble être monophylétique, et ainsi compense l'un des trois domaines de la vie. Les deux autres domaines, bactéries et Archaea, sont procaryotes et ne ont aucune des caractéristiques ci-dessus. Eucaryotes représentent une infime minorité de tous les êtres vivants; même dans un corps humain, il ya 10 fois plus de microbes que de cellules humaines. Toutefois, en raison de leur beaucoup plus grande taille leur biomasse collective dans le monde est estimé à peu près égale à celle de procaryotes.
caractéristiques cellulaires
Les cellules eucaryotes sont généralement beaucoup plus grands que ceux de les procaryotes. Ils ont une variété de membranes et de structures internes, appelées organelles, et un cytosquelette composée de microtubules, microfilaments, et filaments intermédiaires, qui jouent un rôle important dans la définition de l'organisation et de la forme de la cellule. Eucaryote ADN est divisé en plusieurs faisceaux linéaires appelés les chromosomes, qui sont séparées par une broche de microtubules pendant la division nucléaire.

Membrane interne
Cellules eucaryotes comprennent une variété de structures membranaires, collectivement dénommés endomembranaire système. Compartiments simples, appelés ou vésicules vacuoles, peuvent se former par bourgeonnement autres membranes. De nombreuses cellules ingèrent la nourriture et d'autres matériaux à travers un processus de endocytose, où la membrane externe invagine puis pince off pour former une vésicule. Il est probable que la plupart des autres organelles liées à la membrane sont en fin de compte dérivés de ces vésicules.
Le noyau est entouré d'une double membrane (communément appelée enveloppe nucléaire), avec des pores qui permettent matière à se déplacer dans et hors. Diverses extensions tubeless et semblables à des feuilles de la forme de membrane nucléaire ce qu'on appelle la réticulum endoplasmique ou ER, qui est impliqué dans le transport des protéines et de la maturation. Il comprend le RE rugueux où ribosomes sont attachés à synthétiser des protéines, qui entrent dans l'espace intérieur ou de la lumière. Par la suite, ils entrent généralement vésicules qui bourgeonnent hors du RE lisse. Dans la plupart des eucaryotes, ces vésicules de protéines porteurs sont libérés et encore modifiés en piles de vésicules aplaties, appelés Organismes ou dictyosomes Golgi.
Les vésicules peuvent être spécialisées à des fins diverses. Par exemple, lysosomes contiennent des enzymes qui décomposent le contenu des vacuoles alimentaires, et peroxysomes sont utilisés pour briser le peroxyde, qui est toxique autrement. Beaucoup protozoaires ont vacuoles contractiles, qui collectent et expulser l'excès d'eau, et extrusomes, qui expulsent matériau utilisé pour détourner les prédateurs ou la capture des proies. Chez les plantes supérieures, la plupart du volume d'une cellule est repris par une vacuole centrale, qui maintient sa pression osmotique principalement.

1) Membrane interne
2) Membrane externe
3) Crista
4) Matrice
Les mitochondries et les plastes
Les mitochondries sont des organites présents dans presque tous les eucaryotes. Ils sont entourés de deux membranes (chacun un bicouche phospholipidique), l'intérieur de laquelle est pliée en invaginations appelées crêtes, où respiration aérobie a lieu. Les mitochondries contiennent leur propre ADN. Ils sont maintenant généralement considérés comme ayant développé à partir de procaryotes endosymbiotiques, probablement protéobactéries. Les quelques protozoaires qui manquent mitochondries ont été trouvés pour contenir organites mitochondrie dérivés, tels que hydrogénosomes et mitosomes; et donc probablement perdu les mitochondries secondairement.
Les plantes et les différents groupes d' algues ont aussi plastes. Encore une fois, ceux-ci ont leur propre ADN et élaborés à partir de endosymbiotes, dans ce cas, cyanobactéries. Ils prennent généralement la forme de chloroplastes, qui, comme les cyanobactéries contiennent chlorophylle et produire des composés organiques (tels que glucose ) par photosynthèse . D'autres sont impliqués dans la conservation des aliments. Bien que plastes ont probablement eu une origine unique, tous les groupes ne contenant plastidiales sont étroitement liés. Au lieu de cela, certains eucaryotes ont eux obtenu à partir d'autres par endosymbiose secondaire ou l'ingestion.
Endosymbiotiques origines ont également été proposés pour le noyau, pour lequel voir ci-dessous, et pour eucaryote flagelles, censé avoir développé à partir de spirochètes. Ce ne est généralement pas accepté, à la fois d'un manque de preuve cytologique et la difficulté à concilier cela avec la reproduction cellulaire.
Structures du cytosquelette
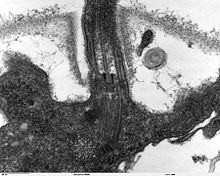
De nombreux eucaryotes ont des projections cytoplasmiques mobiles longs et minces, appelé flagelles, ou semblables structures appelées cils. Flagelles et cils sont parfois appelés undulipodia, et sont diversement impliqués dans le mouvement, l'alimentation, et la sensation. Elles sont composées essentiellement de tubuline. Ce sont entièrement distincte de flagelles procaryotes. Ils sont pris en charge par un faisceau de microtubules provenant d'une basale du corps, également appelé cinétosome ou centriole, caractéristique disposé comme neuf doublets entourant deux maillots. Flagelles peut également avoir des poils, ou mastigonèmes et échelles reliant les membranes et les tiges internes. Leur intérieur est continue avec la cellule de cytoplasme.
Microfilamental structures composées par actine et des protéines de liaison d'actine, par exemple, α- actinine, fimbrine, filamine sont présents dans les couches corticales et faisceaux submembraneous, ainsi. protéines de moteur de microtubules, par exemple, dynéine ou kinésine et l'actine, par exemple, myosines fournissent caractère dynamique du réseau.
Les centrioles sont souvent présents même dans des cellules et des groupes qui ne ont pas de flagelles. Ils se produisent généralement dans les groupes d'un ou de deux, appelée kinetids, qui donnent lieu à diverses racines microtubulaires. Ceux-ci forment une composante principale de la structure du cytosquelette, et sont souvent assemblés au cours de plusieurs divisions cellulaires, avec un flagelle retenu de la mère et une autre dérive. Les centrioles peut également être associée à la formation d'une broche lors de la division nucléaire.
Importance des structures du cytosquelette est souligné dans la détermination de la forme des cellules, ainsi que leurs composants essentiels étant de réponses migrateurs comme chimiotactisme et chimiokinèse. Certains protistes ont divers autres organites des microtubules soutenu. Ceux-ci comprennent le radiolaires et Heliozoa, qui produisent axopodia utilisé dans flottation ou pour capturer leurs proies, et la haptophytes, qui ont un organite flagelle comme particulière appelée haptonema.
la paroi cellulaire
Les cellules de plantes, les champignons et la plupart des Chromalveolates ont une paroi cellulaire, une couche relativement rigide à l'extérieur du membrane cellulaire, la cellule fournissant un support structurel à, la protection, et un mécanisme de filtrage. La paroi cellulaire empêche également une expansion excessive lorsque l'eau pénètre dans la cellule.
Chez les plantes, le principal polysaccharides constituant les parois cellulaires primaires de plantes terrestres sont cellulose, hémicellulose, et la pectine. La cellulose microfibrilles sont liés par l'intermédiaire d'attaches hémicellulosiques pour former le réseau de l'hémicellulose de la cellulose, qui est noyée dans la matrice de pectine. L'hémicellulose la plus courante dans la paroi cellulaire primaire est xyloglucane.
Les différences entre les cellules eucaryotes
Il ya beaucoup de différents types de cellules eucaryotes, si les animaux et les plantes sont les eucaryotes les plus familiers, et fournissent donc un excellent point de départ pour comprendre la structure eucaryote. Les champignons et les nombreux protistes ont quelques différences importantes, cependant.
cellule animale
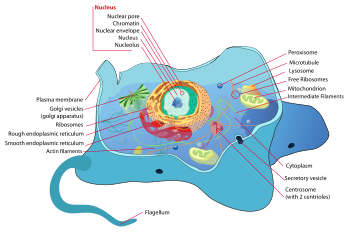

Une cellule animale est une forme de cellule eucaryote qui fait beaucoup tissus animaux . La cellule animale est distinct des autres eucaryotes, et plus particulièrement des cellules végétales , comme ils manquent les parois des cellules et chloroplastes. Ils ont aussi plus petit vacuoles. En raison de l'absence d'un rigide la paroi cellulaire, des cellules animales peut adopter une variété de formes. Un cellule phagocytaire peut même engloutir d'autres structures.
Il ya beaucoup de différents types de cellules. Par exemple, il existe environ 210 types de cellules distinctes dans le corps humain adulte.
Cellule végétale
Plante cellules sont tout à fait différente de celle des cellules des autres organismes eucaryotes. Leurs caractéristiques sont:
- Un grand central vacuole (fermé par une membrane, la tonoplaste), qui maintient la cellule de turgescence et contrôle le mouvement des molécules entre le cytosol et sève
- Un primaire la paroi cellulaire contenant cellulose, hémicellulose et pectine, déposé par la protoplastes à l'extérieur de la membrane cellulaire; Cela contraste avec la parois cellulaires des champignons , qui contiennent la chitine, et la enveloppes cellulaires des procaryotes, dans lequel peptidoglycanes sont les principales molécules structurelles
- Le plasmodesmes, reliant pores dans la paroi cellulaire qui permettent à chaque cellule végétale pour communiquer avec d'autres cellules adjacentes; ce est différent du système fonctionnellement analogue de jonctions communicantes entre les cellules animales.
- Plastids, en particulier chloroplastes contenant chlorophylle, le pigment qui donne plantes leur couleur verte et leur permet d'effectuer la photosynthèse
- Les plantes supérieures, y compris conifères et plantes à fleurs (angiospermes) ne ont pas la flagelles et centrioles qui sont présents dans les cellules animales
La cellule fongique

1- 2- hyphes mur septum 3- 4- mitochondrie Vacuole 5- cristal Ergostérol 6- ribosome 7- 8- réticulum endoplasmique Nucleus corps 9- 10- membrane lipidique plasma 11- Appareil Spitzenkörper 12 Golgi
Les cellules fongiques sont les plus semblables à des cellules animales, avec les exceptions suivantes:
- Une paroi de cellule qui contient chitine
- Moins définition entre les cellules; la hyphes de champignons supérieurs ont des parois poreuses appelé des cloisons, qui permettent le passage du cytoplasme, des organelles, et, parfois, des noyaux. Champignons primitifs ont peu ou pas de cloisons, de sorte que chaque organisme est essentiellement une supercellule multinucléé géant; Ces champignons sont décrits comme coenocytic.
- Seule la plus primitive champignons, chytrides, ont flagelles.
D'autres cellules eucaryotes
Les eucaryotes sont un groupe très diversifié, et leurs structures cellulaires sont également diverses. Beaucoup ont des parois cellulaires; beaucoup ne ont pas. Beaucoup ont chloroplastes, dérivés de l'endosymbiose primaire, secondaire, voire tertiaire; et beaucoup le font pas. Certains groupes ont des structures uniques, tels que la cyanelles du glaucophytes, la haptonema du haptophytes, ou la ejectisomes du cryptomonades. D'autres structures, comme pseudopodes, se trouvent dans divers groupes eucaryotes sous différentes formes, comme le lobose amoebozoans ou l'Reticulose foraminifères.
Reproduction
Division nucléaire est souvent coordonnée avec la division cellulaire. Ceci a lieu généralement par mitose, un processus qui permet à chaque noyau fille pour recevoir un exemplaire de chaque chromosome. Dans la plupart des eucaryotes, il existe également un procédé de reproduction sexuée, impliquant généralement une alternance entre générations haploïdes, dans lequel une seule copie de chaque chromosome est présent, et générations diploïdes, dans lequel deux sont présents, se produisant par la fusion nucléaire (syngamie) et méiose. Il existe des variations considérables dans ce modèle, cependant.
Les eucaryotes ont une surface plus petite par rapport au volume que les procaryotes, et ont donc des taux métaboliques inférieurs et de plus longs temps de génération. Dans certains organismes pluricellulaires, des cellules spécialisées pour le métabolisme auront élargie des surfaces, telles que Vili intestinaux.
Classification

Même retour à l'Antiquité les deux clades de animaux et plantes ont été reconnus. Ils ont eu la rang taxonomique des Uni (biologie) par Linné . Bien qu'il comprenait les champignons avec des plantes avec quelques réserves, il a réalisé plus tard qu'ils sont tout à fait distincte et justifient un royaume séparé, dont la composition ne était pas tout à fait clair jusqu'à ce que les années 1980. Les divers eucaryotes unicellulaires ont été placés à l'origine avec des plantes ou des animaux quand ils sont devenus connus. Le biologiste allemand Georg A. Goldfuss inventé le mot protozoaires en 1830 de se référer à des organismes tels que ciliés et coraux, et ce groupe a été élargi jusqu'à ce qu'il englobe tous les eucaryotes unicellulaires, et compte tenu de leur propre royaume, le Protistes par Ernst Haeckel en 1866. Les eucaryotes ainsi venu à être composé de quatre royaumes:
- Royaume Protistes
- Uni Plantae
- Uni champignons
- Uni Animalia
Les protistes ont été comprises comme des «formes primitives", et donc une année évolutif, unis par leur nature unicellulaire primitive. Le désenchevêtrement des divisions profondes dans la arbre de vie se est seulement vraiment aller avec le séquençage de l'ADN, conduisant à un système de domaines plutôt que royaumes que de niveau supérieur rang mises de l'avant par Carl Woese, unissant tous les royaumes eucaryotes sous le domaine eucaryote. Dans le même temps, travailler sur l'arbre de protistes intensifiée, et est toujours activement aujourd'hui. Plusieurs autres classifications ont été transmis, mais il n'y a pas de consensus dans le domaine.
Un classement produite en 2005 pour la Société internationale de Protistologists, qui reflète le consensus du temps, divisé les eucaryotes en six «supergroupes» prétendument monophylétiques. Bien que le classement publié délibérément n'a pas utilisé les rangs taxonomiques formelles, d'autres sources ont traité chacun des six comme un royaume séparé.
| Excavata | Divers protozoaires flagellés |
| Amoebozoa | La plupart lobose amoeboids et myxomycètes |
| Opisthokonta | Animaux , champignons , choanoflagellés, etc. |
| Rhizaria | Foraminifères, Radiolaires et divers autres protozoaires amiboïdes |
| Chromalveolata | Straménopiles (ou Heterokonta), Haptophyta, Cryptophyta (ou cryptomonades), et Alveolata |
| Archaeplastida (ou Primoplantae) | Les plantes terrestres, algues vertes, algues rouges, et glaucophytes |
Cependant, dans la même année (2005), des doutes ont été exprimés quant à savoir si certains de ces supergroupes étaient monophylétique, en particulier le Chromalveolata, et un examen en 2006 a noté le manque de preuves pour plusieurs des prétendus six supergroupes.
Phylogénie
arbres ARNr construits durant les années 1980 et 1990 ont laissé la plupart des eucaryotes dans un groupe non résolue "couronne" (pas techniquement un véritable couronne), qui était habituellement divisée par la forme de la crêtes mitochondriales; voir eucaryotes couronne. Les quelques groupes que le manque mitochondries ramifié séparément, et ainsi de l'absence était considérée comme primitive; mais ce est maintenant considéré comme un artéfact long branche attraction, et ils sont connus pour leur ont perdu secondairement.
En 2011, il est largement admis que le Rhizaria appartiennent à la straménopiles et Alveolata, dans un clade surnommé le Harosa, de sorte que Rhizara ne est pas l'un des principaux groupes d'eucaryotes; également que le Amoeboza et Opisthokonta sont chacun monophylétique et forment un clade, souvent appelé le unikonts. Au-delà de cela, il ne semble pas y avoir un consensus.
Il a été estimé qu'il pourrait y avoir 75 lignées distinctes des eucaryotes. La plupart de ces lignées sont protistes.
Les tailles de génome eucaryote connus varient de 8,2 mégabases (MB) Babesia bovis à 112,000-220,050 Mb dans la dinoflagellés Prorocentrum micans ce qui suggère que le génome de la cellule eucaryote ancestrale a subi des variations considérables au cours de son évolution. Le dernier ancêtre commun de tous les eucaryotes est soupçonné d'avoir été un protiste phagotrophic avec un noyau, au moins un centriole et cil, les mitochondries aérobie facultative, le sexe ( méiose et syngamie), un sommeil kyste avec une paroi de cellule de la chitine et / ou la cellulose et peroxysomes. Plus tard endosymbiose conduit à la propagation de plastes dans certains lignages.
Chromalveolata + Rhizaria
Certaines analyses démonter l'Chromalveolata + Rhizaria, montrant des relations étroites avec l'Archaeplastida. Par exemple, en 2007, Burki et al. A produit un arbre de la forme ci-dessous.
| |||||||||||||||||||||||||||||||||||||||||||
Bikonts et unikonts
Dans une autre analyse, le Hacrobia sont présentés imbriqué dans l'Archaeplastida, qui forment ensemble un clade avec la plupart des Excavata, avant de rejoindre le clade SAR de straménopiles, Alveolata et Rhizaria. Ensemble, tous ces groupes constituent le bikonts, Amoebozoa et la formation de la Opisthokonta unikonts.
| ||||||||||||||||||||||||||||||||||||||||
La division des eucaryotes en deux clades primaires, unikonts et bikonts, dérivé d'un organisme uniflagellar ancestrale et un organisme biflagellar ancestrale, respectivement, avait été suggéré plus tôt.
Une étude 2012 a produit une division quelque peu similaire, tout en notant que les termes "unikonts" et "bikonts" ne ont pas été utilisés dans le sens originel.
| eucaryotes |
| ||||||||||||||||||||||||||||||
Expanded Chromalveolata
D'autres analyses placent le supergroupe SAR dans un Chromalveolata élargi, même si elles diffèrent sur le placement des cinq groupes qui en résultent. Rogozin et al. En 2009 a produit l'arbre ci-dessous, où le cycle primaire est entre le Archaeplastida et tous les autres eucaryotes.
| |||||||||||||||||||||||||||||||||||||
Plus couramment l'Chromalveolata élargi est présenté comme plus étroitement liée à la Archaeplastida, produire un arbre de la forme ci-dessous.
| |||||||||||||||||||||||||||||||||||||
Vues alternatifs
Un article publié en 2009 qui re-examiné les données utilisées dans certaines des analyses présentées ci-dessus ainsi que l'exécution de nouvelles, fortement suggéré que le Archaeplastida sont polyphylétique. La phylogénie enfin proposé dans le papier est illustré ci-dessous.
| eucaryotes |
| |||||||||||||||||||||||||||||||||
Il ya aussi des petits groupes d'eucaryotes - y compris le genre Collodictyon, la telonemids et biliphytes - dont la position est incertaine ou semble tomber en dehors des grands groupes. Globalement, il semble que si des progrès ont été réalisés, il ya encore des incertitudes très importantes dans l'histoire de l'évolution et de la classification des eucaryotes. Comme l'a dit Roger & Simpson en 2009 "avec le rythme actuel de changement dans notre compréhension de l'arbre de la vie eucaryote, nous devons procéder avec prudence."
Origine des eucaryotes


L'origine de la cellule eucaryote est considéré comme une étape importante dans l'évolution de la vie, car ils comprennent toutes les cellules complexes et presque tous les organismes multicellulaires. Le calendrier de cette série d'événements est difficile à déterminer; Knoll (2006) suggère qu'ils ont développé il ya environ 1,6 à 2100000000 années. Certains acritarches sont connus d'au moins 1650 millions d'années, et l'algue possible Grypania a été trouvé aussi loin que il ya 2,1 milliards années.
Structures organisées de vie ont été trouvés dans schistes noirs de la Formation paléoprotérozoïque Francevillian B au Gabon, datés à l'âge de 2,1 milliards années. La vie eucaryote pourrait avoir évolué à cette époque. Fossiles qui sont clairement liés à des groupes modernes commencent à apparaître environ 1,2 milliards d'années, sous la forme d'un algue rouge, bien que des travaux récents suggèrent l'existence d'fossilisé algues filamenteuses dans le Vindhya bassin datant peut-être il ya 1,6 à 1700000000 ans.
Biomarkers suggèrent qu'au moins souches eucaryotes surgi même plus tôt. La présence de stéranes en Australie schistes indique que les eucaryotes étaient présents dans ces roches datées de vieilles de 2,7 milliards d'années.
Relations avec les Archaea
Les eucaryotes sont plus étroitement liés à Archaea de bactéries , au moins en termes d'ADN nucléaire et machinerie génétique, et une idée controversée est de les placer avec Archaea dans le clade Neomura. Cependant, à d'autres égards, comme la composition de la membrane, les eucaryotes sont similaires aux bactéries. Trois principales explications de cette ont été proposées:
- Les eucaryotes résulte de la fusion complète de deux ou plusieurs cellules, dans lequel le cytoplasme formé à partir d'une eubactérie, et le noyau à partir d'un archaeon, d'un virus, ou à partir d'un pré-cellule.
- Eucaryotes développés à partir Archaea, et ont acquis leurs caractéristiques eubactériennes de la proto-mitochondrie.
- Eucaryotes et Archaea développés séparément à partir d'un eubactérie modifié.
Il existe également la théorie Kronocyte de l'origine de la cellule eucaryote. Ce postule qu'une cellule eucaryote primitive émergé du monde pré-ADN, mais a conservé la chimie à base d'ARN-tôt à partir de laquelle toute vie moderne a émergé. Cette cellule primitive est appelé Kronocyte. Selon cette hypothèse un Kronocyte base-ARN coexistait avec les Archaea à base d'ADN (et probablement eubactéries) et est devenu la cellule eucaryote moderne après un certain nombre de grands endosymbioses-le premier a été l'incorporation d'une Archaea qui a introduit métabolisme de l'ADN et le noyau, puis l'incorporation d'un alphaproteobacter qui est devenu le mitochondries (et les bactéries photosynthétiques trouvés dans les usines d'aujourd'hui comme les chloroplastes). L'hypothèse Kronocyte explique le grand nombre de gènes qui sont aujourd'hui seulement constaté chez les eucaryotes mais pas chez les Archaea ou bactéries.
Système de mitochondries et Endomembrane
Les origines du système endomembranaire et les mitochondries ne sont pas claires. L'hypothèse phagotrophic propose que les membranes de type eucaryote qui ne ont pas de paroi cellulaire proviennent d'abord, avec le développement de l'endocytose, alors que les mitochondries ont été acquis par l'ingestion comme endosymbionts. L'hypothèse syntrophique propose que le proto-eucaryote se est appuyé sur le proto-mitochondrie pour la nourriture, et ainsi finalement grandi pour entourer. Voici les membranes origine après l'engloutissement de la mitochondrie, en partie grâce à gènes mitochondriaux (la hypothèse de l'hydrogène est une version particulière).
Dans une étude utilisant des génomes de construire supertrees, Pisani et al. (2007) suggèrent que, avec la preuve qu'il n'y a jamais un eucaryote de mitochondrie-moins, les eucaryotes ont évolué à partir d'un syntrophie entre un archées étroitement liée à Et un Thermoplasmatales α-protéobactérie, probablement un symbiose entraînée par le soufre ou de l'hydrogène. La mitochondrie et son génome est un vestige de la endosymbiote de α-protéobactéries.
Les hypothèses pour l'origine des eucaryotes
Différent hypothèses ont été proposées sur la façon dont les cellules eucaryotes est entré en existence. Ces hypothèses peuvent être classés en deux catégories distinctes - modèles autogènes et modèles chimériques.
Modèles autogènes
 |
| Un modèle pour l'origine autogène des eucaryotes. |
Autogènes modèles proposent que l'origine une cellule de proto-eucaryote contenant un noyau d'abord existé, et plus tard acquis mitochondries . Selon ce modèle, une grande procaryote développé invaginations dans son membrane de plasma de façon à obtenir suffisamment la superficie totale pour le service de sa le volume cytoplasmique. Comme les invaginations différenciée en fonction, certains sont devenus séparés compartiments-donnant lieu à la endomembrane système, y compris la reticulum endoplasmique, appareil de Golgi, la membrane nucléaire et la membrane unique structures telles que lysosomes. Les mitochondries sont proposées à venir de la endosymbiose d'un aérobie protéobactérie, et il est supposé que toutes les lignées eucaryotes qui ne ont pas acquis les mitochondries se sont éteints. Les chloroplastes sont venus à propos d'un autre événement impliquant endosymbiotique cyanobactéries. Depuis tous les eucaryotes ont mitochondries, mais tous ne ont pas les chloroplastes, les mitochondries sont pensé avoir venir en premier. Ceci est le la théorie de l'endosymbiose série.
Certains modèles proposent que les origines de la double couche organites tels que les mitochondries et chloroplastes dans la cellule eucaryote proto-est due au cloisonnement des ADN des vésicules qui ont été formés à partir des invaginations secondaires ou des replis plus détaillées de la membrane cellulaire.
Modèles chimériques
Modèles chimériques affirment que deux cellules procaryotes existaient initialement - un archaeon et un bactérie . Ces cellules ont subi un processus de fusion soit par une fusion physique ou par endosymbiose, ce qui conduit à la formation d'une cellule eucaryote. Dans ces modèles chimériques, certaines études affirment en outre que les mitochondries origine d'un ancêtre bactérien tandis que d'autres mettent l'accent sur le rôle des processus endosymbiotiques derrière l'origine des mitochondries.
Selon le procédé de symbiose mutualiste, les hypothèses peuvent être classés comme - la série théorie endosymbiotique (SET), le hypothèse de l'hydrogène (la plupart du temps un processus de symbiose où le transfert d'hydrogène a lieu entre les différentes espèces), et l'hypothèse de syntrophie.
Selon la théorie endosymbiotique série, une union entre un mobiles bactérie anaérobie (comme Spirochaeta) et un crenarchaeon thermoacidophile (comme Thermoplasma qui est sulfurogènes dans la nature) ont donné lieu à la présente eucaryotes de jours. Cette union a créé un organisme mobile capable de vivre dans les eaux acides et sulfureuses déjà existants. L'oxygène est connue pour sa toxicité pour les organismes qui ne ont pas le nécessaire machinerie métabolique. Ainsi, le archaeon fourni la bactérie avec un environnement très bénéfique réduite (soufre et le sulfate ont été réduits en sulfure). En microaérophilie, l'oxygène a été réduit à l'eau créant ainsi une plate-forme de bénéfice mutuel. La bactérie, d'autre part, la nécessaire contribué produits de fermentation et électrons accepteurs ainsi que sa fonction de motilité à l'archée gagnant ainsi une piscine la motilité de l'organisme. D'un consortium d'ADN bactérien et Archaea origine du nucléaire le génome des cellules eucaryotes. Spirochètes ont donné lieu à des caractéristiques mobiles de cellules eucaryotes. Unifications endosymbiotiques des ancêtres des alpha-protéobactéries et les cyanobactéries, ont conduit à l'origine des mitochondries et des plastides respectivement. Par exemple, Thiodendron a été connu pour avoir son origine par l'intermédiaire d'un ectosymbiotic processus fondé sur une même syntrophie du soufre existant entre les deux types de bactéries - Desulphobacter et Spirochaeta. Cependant, une telle association basée sur la symbiose mobiles ne ont jamais été observée pratiquement. Aussi il ne existe aucune preuve de archaeans et spirochètes adapter à des environnements à base d'acide intenses.
Dans l'hypothèse d'hydrogène, la liaison d'un symbiotique anaérobie et autotrophes archaeon méthanogène (hôte) avec une alpha-protéobactérie (le symbiote) a donné lieu à des eucaryotes. L'hôte utilisé un atome d'hydrogène (H 2) et du dioxyde de carbone (CO 2) pour produire du méthane tandis que le symbiont, apte à la respiration aérobie, expulsé H 2 et de CO 2 en tant que sous-produits du processus de fermentation anaérobie. Méthanogène environnement de l'hôte a travaillé comme un puits de H 2, qui a abouti à la fermentation bactérienne accrue. Transfert de gènes endosymbiotique (EGT) a agi comme un catalyseur pour l'hôte d'acquérir des symbiotes glucides et le métabolisme tour hétérotrophe dans la nature. Par la suite, le méthane formant la capacité de l'hôte a été perdue. Ainsi, les origines de l'organite hétérotrophes (symbiote) sont identiques aux origines de la eucaryote lignée. Dans cette hypothèse, la présence de H 2 représente la force sélective qui a forgé les eucaryotes de procaryotes.
L'hypothèse de syntrophie est développé contrairement à l'hypothèse d'hydrogène et propose l'existence de deux événements symbiotiques. Selon cette théorie, eukaryogenesis (c. origine des cellules eucaryotes) se est produite basée sur la symbiose métabolique (syntrophie) entre une archée méthanogène et un delta-protéobactérie. Cette symbiose syntrophique a d'abord été facilitée par H 2 transfert entre espèces différentes dans des environnements anaérobies. Dans les premiers stades, un alpha-protéobactérie est devenu membre de cette intégration, et développé plus tard dans la mitochondrie. Le transfert de gènes à partir d'un delta-protéobactérie à un archaeon conduit à l'archée méthanogène développer dans un noyau. Le archaeon constitué l'appareil génétique tandis que le delta-protéobactérie contribué à la caractéristiques cytoplasmiques. Cette théorie comporte deux forces de sélection qui ont été nécessaires pour être considéré à l'époque du noyau évolution - (a) présence de partitionnement métabolique afin d'éviter les effets néfastes de la co-existence de et anabolique voies cellulaires cataboliques, et (b) la prévention de anormale biosynthèse des protéines qui se produisent en raison d'une vaste propagation de introns dans les gènes d'archées après l'acquisition de la mitochondrie et la perte de méthanogénèse.
Ainsi, l'origine des eucaryotes par des procédés endosymbiotiques a été largement reconnu et accepté jusqu'à présent. Les mitochondries et les plastes ont été connus pour provenir d'un ancêtre bactérien durant adaptation parallèle à anaérobiose. Cependant, il reste encore un plus grand besoin pour évaluer la question de savoir combien la complexité eucaryote qui est originaire via une mise en œuvre de ces symbiogenetic théories.