
Ptérosaure
Contexte des écoles Wikipédia
Arrangeant une sélection Wikipedia pour les écoles dans le monde en développement sans internet a été une initiative de SOS Enfants. parrainage SOS enfant est cool!
| Ptérosaures Plage temporelle: Late Trias Crétacé supérieur, 210-65.5Ma | |
|---|---|
 | |
| Réplique Les squelettes de Geosternbergia, femelle (à gauche) et mâle (à droite) | |
| Classification scientifique | |
| Uni: | Animalia |
| Embranchement: | Chordata |
| Classe: | Reptilia |
| Clade: | Ornithodira |
| Ordre: | † Pterosauria Kaup, 1834 |
| Suborders | |
† Pterodactyloidea | |
| Synonymes | |
| |
Ptérosaures (pron .: / t ɛr ɵ s ɔr /, Du grec πτερόσαυρος, pterosauros, ce qui signifie «lézard ailé") volaient reptiles de la clade ou commander Pterosauria. Ils ont existé de la fin du Trias à la fin des Crétacé Période (220-65 Il ya des millions d'années). Ptérosaures sont les premiers vertébrés connus pour avoir évolué vol motorisé. Les ailes sont formées par une membrane de peau, les muscles et d'autres tissus se étendant des chevilles à un quatrième doigt considérablement allongée. Les premières espèces eu, mâchoires dentées entièrement longues et longues queues, tandis que les formes ultérieures avaient une queue très réduit, et certains ne avaient pas les dents. Beaucoup de manteaux à fourrure arboré constitués de filaments ressemblant à des cheveux dits pycnofibres , qui couvraient leurs corps et parties de leurs ailes. Ptérosaures porté sur une vaste gamme de tailles adultes, de la très petite Nemicolopterus aux plus grandes créatures volantes connues de tous les temps, y compris Quetzalcoatlus et Hatzegopteryx.
Ptérosaures sont souvent mentionnés dans les médias populaires et par le grand public comme battant dinosaures , mais ce est incorrect. Le terme «dinosaure» est correctement restreint aux seuls reptiles descendus à partir du dernier ancêtre commun des groupes Saurischia et Ornithischia (clade Dinosauria, qui comprend les oiseaux ), et le consensus scientifique actuel est que ce groupe exclut les ptérosaures, ainsi que les différents groupes de éteinte reptiles marins, tels que les ichtyosaures , plésiosaures, et mosasaures. Ptérosaures sont également qualifiées, à tort ptérodactyles, en particulier par des journalistes. "Pterodactyl» se réfère spécifiquement aux membres du genre Pterodactylus, et plus largement pour les membres du sous-ordre Pterodactyloidea.
Description
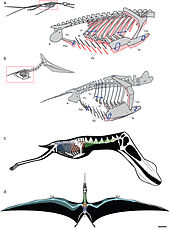
L'anatomie de ptérosaures a été très modifiée de leurs ancêtres reptiliens pour les demandes de vol. Ptérosaure os étaient creux et rempli d'air, comme les os d' oiseaux . Ils avaient un Keeled sternum qui a été développé pour la fixation de vol muscles et un élargie cerveau qui montre des fonctions spécialisées associés au vol. Dans certains ptérosaures plus tard, l'épine dorsale sur les épaules fusionné dans une structure connue comme Notarium, qui a servi à rigidifier le torse pendant le vol, et de fournir un support stable pour le omoplate (omoplate).
Coulisses
ailes Pterosaur ont été formés par des membranes de la peau et d'autres tissus. Les membranes primaires attachés à la quatrième extrêmement longue chaque doigt de armer et étendu le long des côtés du corps jusqu'aux chevilles.
Si, historiquement, pensé comme de simples structures composées, coriaces de la peau, la recherche a montré depuis que les membranes des ailes de ptérosaures étaient des structures effectivement très complexes et dynamiques adaptés à un style actif de vol. Les ailes extérieures (de la pointe du coude) ont été renforcés par des fibres rapprochées appelés actinofibrils. Les actinofibrils eux-mêmes composés de trois couches distinctes de l'aile, formant un empilage croisé lorsque superposée à une autre. La fonction réelle des actinofibrils est inconnue, comme ce est le matériau exacte à partir de laquelle ils ont été faits. En fonction de leur composition exacte (de la kératine, les muscles, les structures élastiques, etc.), ils ont peut-être été de renforcement ou renforcement des agents dans la partie extérieure de l'aile. Les membranes des ailes contenaient également une fine couche de muscle, le tissu fibreux, et un système circulatoire unique complexe de boucle vaisseaux sanguins.
Comme en témoignent les cavités creusées dans les os des ailes de grandes espèces et les tissus mous conservés dans au moins un échantillon, certains ptérosaures étendre leur système de sacs aériens respiratoires (voir la section de paléobiologie ci-dessous) dans la membrane de l'aile elle-même.
Pièces de l'aile de ptérosaure

La membrane de l'aile de ptérosaure est divisé en trois unités de base. Le premier, appelé le propatagium ("première membrane"), était la partie la plus en avant de l'aile et attaché entre le poignet et l'épaule, la création de la "pointe" pendant le vol. Cette membrane peut avoir incorporé les trois premiers doigts de la main, comme en témoignent certains spécimens. Le brachiopatagium ("membrane de bras") était le composant principal de l'aile, se étendant de la quatrième doigt très allongé de la main pour les membres postérieurs (mais où exactement sur les membres postérieurs, il ancrée est controversée et peut avoir varié entre les espèces, voir ci-dessous ). Enfin, au moins certains groupes de pterosaur avaient une membrane qui se étend entre les jambes, peut se connecter à ou incorporant la queue, dite uropatagium; la mesure de cette membrane ne est pas certain, que des études sur Sordes semblent suggérer que simplement connecté les jambes, mais ne implique pas la queue (rendant un cruropatagium). Il est généralement admis que si ptérosaures non-pterodactyloid avaient une plus large uro / cruropatagium, avec pterodactyloids ne ayant membranes courent le long des jambes; Pteranodon en particulier aurait pu développer / réaménagé un uropatagium, étant donné la structure de la queue.
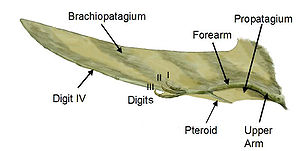
Un os unique de pterosaurs, connu sous le pteroid, relié au poignet et contribué à soutenir une membrane vers l'avant (la propatagium) entre le poignet et l'épaule. Preuve de sangle entre les trois doigts libres de la patte avant de ptérosaure suggère que cette membrane avant peut-être plus étendue que la simple connexion pteroid à épaule traditionnellement représenté dans les restaurations de la vie. La position de l'os se pteroid a été controversée. Certains scientifiques, notamment Matthew Wilkinson, ont fait valoir que le pteroid fait avant, étendant la membrane avant. Toutefois, ce point de vue a été vivement réfuté dans un document de 2007 par Chris Bennett, qui a montré que le pteroid n'a pas articuler comme le pensait précédemment, et ne pouvait pas avoir l'avant pointu, mais plutôt vers l'intérieur vers le corps comme on le pensait traditionnellement.
Trois sources de données, morphologique, de développement et histologique, indiquent que le pteroid est un véritable os, plutôt que le cartilage ossifié. L'origine de la pteroid ne est pas clair: il peut être un carpien modifié, le premier métacarpien, ou neomorph (nouvel os).
Le poignet de ptérosaure se compose de deux intérieure (proximale) et quatre carpes extérieures (distales) (os du poignet), à l'exclusion de l'os pteroid, qui peut lui-même être un carpien distale modifiée. Les carpes proximaux sont fusionnés en un "syncarpal" des spécimens matures, tandis que trois des carpes distales fusionner pour former un syncarpal distale. Le carpien distale restante, appelée ici la carpien médiane, mais qui a également été appelé le carpien latérale, ou pré-axial distal, se articule sur une facette allongée verticalement biconvexe sur la surface antérieure de la syncarpal distale. Le carpien médial porte une fovéa concave profonde qui se ouvre en avant, le ventre et un peu en dedans, dans lequel les articule pteroid.
Il a été l'argument considérable chez les paléontologues quant à savoir si les principales membranes d'aile (de brachiopatagia) attachés aux membres postérieurs, et si oui, où. Fossiles du rhamphorhynchoid Fuliginosités, la anurognathid Jeholopterus, et un de la pterodactyloid Formation Santana semblent démontrer que la membrane de l'aile ne se attachent aux membres postérieurs, au moins chez certaines espèces. Cependant, modernes chauves-souris et écureuils volants montrent des variations considérables dans la mesure de leurs membranes d'aile et il est possible que, comme ces groupes, différentes espèces de ptérosaures avaient des conceptions différentes de l'aile. En effet, l'analyse des proportions ptérosaure des membres montre qu'il y avait une variation considérable, reflétant peut-être une variété de plans d'ailes.
Beaucoup, sinon la totalité, ptérosaures avaient aussi les pieds palmés.
Crâne, des dents et des crêtes

La plupart des crânes de ptérosaures avaient des mâchoires allongées avec une gamme complète de dents en forme d'aiguille. Dans certains cas, fossilisé tissu bec kératinique a été conservé, quoique sous des formes dentées, le bec est petit et limité à des conseils de la mâchoire et ne implique pas les dents. Certaines formes de pointe sont à bec édenté, tel que le pteranodonts et azhdarchids, et avait plus grand, plus vaste et plus becs d'oiseaux ressemblant.
Contrairement à la plupart archosaurs, qui présentent plusieurs ouvertures dans le crâne devant les yeux, dans la pterosaurs pterodactyloid antéorbitaire ouverture et l'ouverture nasale a été fusionnées en une seule grande ouverture, appelé le fenestra nasoantorbital. Ce susceptibles évolué comme une caractéristique de gain de poids pour alléger le crâne pour le vol.
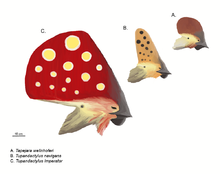
Ptérosaures sont bien connus pour leurs crêtes souvent élaborés. La première et peut-être le plus connu d'entre eux est la crête pointant vers l'arrière distinctif de certains Pteranodon espèces, bien que quelques ptérosaures, comme le tapejarids et Nyctosaurus arborait incroyablement grandes crêtes qui souvent incorporés extensions des tissus mous kératiniques ou d'autres de la base de crête osseuse.
Depuis les années 1990, de nouvelles découvertes et une étude plus approfondie de vieux spécimens ont montré que les crêtes sont beaucoup plus répandue chez les ptérosaures qu'on ne le pensait précédemment, principalement en raison du fait qu'ils ont souvent été prolongés par ou composées entièrement de la kératine, qui ne se fossilise pas aussi souvent comme l'os. Dans les cas de ptérosaures comme Pterorhynchus et Pterodactylus, la véritable ampleur de ces crêtes n'a été découvert en utilisant ultraviolet photographie. La découverte de Pterorynchus et Austriadactylus, deux aigrettes " rhamphorhynchoids ", a montré que même les ptérosaures primitifs avaient crêtes (précédemment, crêtes ont été pensés pour être limité à la plus avancée pterodactyloids).
Pycnofibres
Au moins certains ptérosaures étaient couverts de filaments ressemblant à des cheveux dits pycnofibres, similaire mais pas homologue (partager une structure commune) avec des mammifères cheveux. Pycnofibres cheveux ne étaient pas vrai comme on le voit chez les mammifères, mais une structure unique qui se est développé grâce à une apparence similaire évolution convergente. Bien que dans certains cas actinofibrils (fibres structurelles internes) dans la membrane de l'aile ont été pris pour pycnofibres ou les cheveux vrai, certains fossiles tels que ceux de Sordes pilosus (qui se traduit par «démon poilu") et Jeholopterus ninchengensis faire apparaître les empreintes indubitables de pycnofibres sur la tête et le corps, un peu comme les chauves-souris modernes, un autre exemple d'évolution convergente. On ne sait pas si elles peuvent être homologue à plumes . La présence de pycnofibres (et les exigences de vol) implique que les ptérosaures étaient endothermique (à sang chaud).
Le terme «pycnofibre", ce qui signifie "filament dense", a été inventé dans un document sur les impressions de tissus mous de Jeholopterus par le paléontologue Alexander WA Kellner et collègues en 2009.
Histoire de la découverte
Le premier ptérosaure fossiles a été décrit par le Naturaliste italien Cosimo Collini en 1784. Collini mal interprété son spécimen comme une créature de mer qui a utilisé ses longs membres avant que les palettes. Quelques scientifiques ont continué à soutenir l'interprétation aquatiques, même jusqu'en 1830, quand le zoologiste allemand Johann Georg Wagler suggéré que Pterodactylus utilisé ses ailes comme des nageoires. Georges Cuvier abord suggéré que les ptérosaures volaient créatures en 1801, et a inventé le nom "Ptero-dactyle" en 1809 pour l'échantillon récupéré en Allemagne. Toutefois, en raison de la normalisation des noms scientifiques, le nom officiel de ce genre est devenu Pterodactylus, si le nom "ptérodactyle" a continué à être populaire et mal appliquée à tous les membres de Pterosauria. Les paléontologues évitent aujourd'hui de "ptérodactyle" et préfèrent le terme «ptérosaure". Ils relèguent le terme «ptérodactyle" spécifiquement pour les membres du genre Pterodactylus ou plus largement pour les membres du sous-ordre Pterodactyloidea.

Paléobiologie
Vol

La mécanique du vol de ptérosaure ne sont pas complètement comprises ou modélisées à ce moment.
Katsufumi Sato, un scientifique japonais, a fait des calculs en utilisant les oiseaux modernes et a décidé qu'il était impossible pour un ptérosaure de rester en altitude. Dans le livre posture, la locomotion et paléoécologie des ptérosaures il est théorisé qu'ils étaient capables de voler en raison de l'atmosphère dense riche en oxygène de la Crétacé supérieur. Cependant, il faut noter la fois Katsufumi et les auteurs de la posture, la locomotion et paléoécologie des ptérosaures fondé leur recherche sur les théories désormais obsolètes de ptérosaures étant oiseaux de mer comme, et la limite de taille ne se applique pas aux ptérosaures terrestres comme azhdarchids et tapejarids Par ailleurs, Darren Naish a conclu que les différences atmosphériques entre le présent et le mésozoïque étaient pas nécessaires à la taille géante de ptérosaures.
Cependant, Mark Witton et Mike Habib, de l'Université de Portsmouth et l'Université Johns Hopkins, respectivement, ont fait valoir que les ptérosaures ont utilisé un mécanisme de voûte pour obtenir vol. Une fois dans l'air, les ptérosaures pourraient atteindre des vitesses allant jusqu'à 120 kilomètres par heure (75 mph) et des milliers de kilomètres.
En 1985, la Smithsonian Institution a commandé ingénieur aéronautique Paul MacCready de construire un modèle de travail à mi-échelle de Quetzalcoatlus northropi. La réplique a été lancée avec un treuil au sol. Il a volé à plusieurs reprises en 1986 et a été filmé dans le cadre du film IMAX de la Smithsonian Institution sur l'aile. Cependant, le modèle ne était pas anatomiquement correcte et incarne verticales et horizontales stabilisateurs arrière qui manquaient dans les ptérosaures. La distribution des poids du modèle est aussi différent en raison de la queue plus longue du modèle.
sacs d'air et la respiration
Une étude réalisée en 2009 a montré que pterosaurs avaient un système de sac poumon d'air et une pompe de respiration squelettique contrôlée avec précision, ce qui supporte un modèle de ventilation pulmonaire écoulement dans pterosaurs, analogue à celle des oiseaux. La présence d'un sous-cutanée système de sac d'air dans au moins certains pterodactyloids aurait réduit davantage la densité de l'animal vivant.
Système nerveux
Une étude de ptérosaure cerveau cavités utilisant des rayons X a révélé que les animaux (et Rhamphorhynchus muensteri Anhanguera santanae) avait flocons massif. Le flocculus est une région du cerveau qui intègre les signaux des articulations, muscles, peau et l'équilibre organes.
Les ptérosaures de les flocons occupées 7,5% des animaux de masse du cerveau total, plus que dans tout autre vertébré. Les oiseaux ont exceptionnellement grandes flocons par rapport aux autres animaux, mais ceux-ci ne occupent entre 1 et 2% de la masse totale du cerveau.
Le flocculus envoie des signaux neuronaux qui produisent de petits mouvements automatiques dans les muscles de l'oeil. Ceux-ci gardent l'image sur la rétine d'un animal stable. Ptérosaures ont peut-être eu un si grand flocculus raison de leur taille d'aile grande, ce qui voudrait dire qu'il y avait beaucoup plus l'information sensorielle à traiter.
mouvements de terrain
Les prises de la hanche de ptérosaures sont orientés face légèrement vers le haut, et la tête de la fémur (os de la cuisse) ne est que modérément orientée vers l'intérieur, ce qui suggère que les ptérosaures avaient une position semi-dressées. Il aurait été possible de soulever la cuisse en position horizontale pendant le vol comme les lézards glisse font.
Il y avait un débat considérable si ptérosaures ambulated que ou comme quadrupèdes bipèdes. Dans les années 1980, le paléontologue Kevin Padian a suggéré que les petites ptérosaures avec les membres postérieurs plus tels que Dimorphodon aurait pu marcher ou même de fonctionner bipedally, en plus de voler, comme coureurs sur route. Cependant, un grand nombre de pterosaur trackways ont été retrouvés avec une patte arrière à quatre doigts distinctif et trois orteils pied avant; ce sont les gravures indubitables de ptérosaures marche à quatre pattes.
Contrairement à la plupart des vertébrés, qui marchent sur leurs orteils avec chevilles détenus sur le sol ( digitigrade), empreintes fossiles montrent que les ptérosaures étaient avec l'ensemble du pied en contact avec le sol ( plantigrade), d'une manière similaire à l'homme et l'ours . Empreintes de azhdarchids montrent qu'au moins certains ptérosaures marchaient avec une érection, plutôt que tentaculaire, la posture.

Bien que traditionnellement représenté comme disgracieux et gênant quand sur le terrain, l'anatomie d'au moins certains ptérosaures (en particulier pterodactyloids) suggère qu'ils étaient marcheurs et coureurs compétentes. Les os des membres antérieurs de azhdarchids et ornithocheirids ont été exceptionnellement longue par rapport aux autres ptérosaures, et azhdarchids, les os des bras et des mains (métacarpiens) étaient particulièrement allongée. En outre, azhdarchidé membres antérieurs dans son ensemble ont été dosées de façon similaire à jeûner-course mammifères ongulés. Les membres postérieurs, d'autre part, ne ont pas été conçus pour la vitesse, mais ils ont été comparés avec la plupart longue pterosaurs, et on les laisse pendant une longue longueur de foulée. Alors que les ptérosaures azhdarchidé ne pourraient probablement pas fonctionner, ils auraient été relativement rapide et efficace de l'énergie.
La taille relative des mains et des pieds de ptérosaures (par comparaison avec des animaux tels que les oiseaux modernes) peut indiquer quel type de ptérosaures de style de vie menées sur le terrain. Ptérosaures azhdarchidé avaient relativement petits pieds par rapport à la taille de leur corps et des jambes de longueur, avec la longueur du pied seulement environ 25% -30% de la longueur de la jambe inférieure. Ceci suggère que azhdarchids étaient mieux adaptées à la marche sur un sol sec et relativement solide. Pteranodon avait les pieds légèrement plus grandes (47% de la longueur du tibia), tandis que les ptérosaures filtreurs comme le ctenochasmatoids avait de très grands pieds (69% de la longueur du tibia en Pterodactylus, 84% en Pterodaustro), adapté à la marche dans le sol vaseux, similaire à échassiers modernes.
Les prédateurs naturels
Ptérosaures sont connus pour avoir été mangé par les théropodes . Dans l'édition du 1er Juillet 2004 Nature, paléontologue Eric Buffetaut traite d'un fossile du Crétacé précoce de trois col de l'utérus vertèbres d'un ptérosaure avec la dent cassée d'un spinosaur noyé dans la masse. Les vertèbres sont connu pour ne pas avoir été mangé et exposée à la digestion, comme les joints articulés encore.
Reproduction et de la vie l'histoire

Très peu est connu sur la reproduction de ptérosaure, et les oeufs de ptérosaures sont très rares. L'œuf de ptérosaure abord connu a été trouvé dans les carrières de Liaoning, au même endroit qui a donné les célèbres dinosaures à plumes ''. L'œuf a été écrasé à plat, avec des signes de fissuration, si évidemment les œufs coquilles eu coriaces, comme dans les lézards modernes. Cela a été pris en charge par la description d'un ptérosaure oeuf additionnel appartenant au genre Darwinopterus, décrit en 2011, qui avait aussi une enveloppe de cuir, et, également comme des reptiles modernes mais contrairement oiseaux, était assez faible par rapport à la taille de la mère. Une étude de la structure ptérosaure de coquille d'oeuf et de la chimie publié en 2007 a indiqué qu'il est ptérosaures susceptibles enterrés leurs œufs, comme modernes crocodiles et tortues . Egg-enfouissement aurait été bénéfique à l'évolution précoce des ptérosaures, car elle permet de plus d'adaptations de réduction de poids, mais cette méthode de reproduction aurait également imposer des limites sur la variété des environnements ptérosaures pourraient vivre dans, et peut les avoir désavantagé quand ils ont commencé à faire face à la concurrence écologique des oiseaux .
membranes alaires conservés dans des embryons de ptérosaures sont bien développés, les ptérosaures étaient suggérant prêt à voler peu après la naissance. Les fossiles de ptérosaures seulement quelques jours à une semaine vieux (appelés flaplings) ont été trouvés, ce qui représente plusieurs familles de ptérosaures, y compris pterodactylids, rhamphorhinchids, ctenochasmatids et azhdarchids. Tous préserver os qui montrent un degré relativement élevé de durcissement (sclérose) pour leur âge, et les proportions des ailes semblables aux adultes. En fait, beaucoup de flaplings pterosaur ont été considérés comme des adultes et placés dans des espèces séparées dans le passé. En outre, flaplings se trouvent normalement dans les mêmes sédiments que les adultes et les jeunes de la même espèce, comme les flaplings Pterodactylus et Rhamphorhynchus trouvés dans le Solnhofen calcaire de l'Allemagne, et flaplings Pterodaustro en provenance du Brésil. Toutes se trouvent dans l'environnement aquatique profonde loin de la rive.
On ne sait pas si les ptérosaures pratiquées toute forme de soins parentaux, mais leur capacité à voler dès qu'ils ont émergé de l'œuf et les nombreux flaplings trouvés dans des environnements loin de nids et, comme les adultes a conduit la plupart des chercheurs, y compris Christopher Bennett et David Unwin , de conclure que le jeune étaient dépendants de leurs parents pour une période relativement courte de temps, au cours d'une période de croissance rapide tandis que les ailes ont augmenté assez longtemps pour voler, et ont ensuite quitté le nid à eux-mêmes, peut-être quelques jours de l'éclosion. Alternativement, ils peuvent ont utilisé les produits de jaune stockées pour la nourriture au cours de leurs premiers jours de vie, comme dans les reptiles modernes, plutôt que de dépendre parents pour la nourriture.
taux de ptérosaures de croissance une fois qu'ils ont éclos variée dans les différents groupes. En plus primitif, ptérosaures à longue queue (" rhamphorhynchoids ") tels que Rhamphorhynchus, le taux de croissance moyen au cours de la première année de vie était de 130% à 173%, légèrement plus rapide que le taux de croissance de alligators . La croissance de ces espèces ralenti après la maturité sexuelle, et il aurait fallu plus de trois Rhamphorhynchus ans pour atteindre à la taille maximale. En revanche, la plus avancée, large ptérosaures pterodactyloid tels que Pteranodon a grandi à la taille adulte dans la première année de vie. En outre, pterodactyloids avaient croissance déterminée, ce qui signifie que les animaux ont atteint une taille maximale fixe des adultes et ont cessé de croître.
Modèles d'activité quotidiens
Les comparaisons entre le anneaux sclérales de ptérosaures et les oiseaux et les reptiles modernes ont été utilisées pour déduire les modèles d'activité quotidienne des ptérosaures. Les genres de ptérosaure Pterodactylus, Scaphognathus, et Tupuxuara ont été déduite être diurne, Ctenochasma, Pterodaustro, et Rhamphorhynchus ont été déduite être nocturne, et Tapejara a été déduite d'être cathemeral, être actif toute la journée pour de courts intervalles. En conséquence, le poisson peut manger Ctenochasma et Rhamphorhynchus peut avoir eu des motifs d'activité similaires aux oiseaux de mer nocturnes modernes, et le filtre-alimentation Pterodaustro peut avoir eu des motifs d'activité similaires au moderne les oiseaux qui se nourrissent de ansériforme nuit. Les différences entre les modèles de l'activité Solnhofen ptérosaures Ctenochasma, Rhamphorhynchus, Scaphognathus et Pterodactylus peut également indiquer la séparation des niches entre ces genres.
Evolution et l'extinction
Origines

Parce ptérosaure anatomie a été si fortement modifiée pour le vol, et de ses prédécesseurs "de chaînon manquant" immédiats ne ont pas jusqu'ici été décrit, l'ascendance de ptérosaures ne est pas bien comprise. Plusieurs hypothèses ont été avancées, y compris des liens vers ornithodirans comme Scleromochlus, une ascendance parmi les archosauriformes basales comme Euparkeria, ou parmi les Prolacertiformes (qui comprennent les formes de glisse comme Sharovipteryx).
Deux chercheurs, Chris Bennett (1996) et David Peters (2000), ont trouvé ptérosaures être Prolacertiformes ou étroitement liés à eux. Peters utilise une technique appelée DGS, qui consiste à appliquer les fonctions numériques de recherches de logiciel de retouche photo pour les images de fossiles de ptérosaures. Bennett ne récupéré ptérosaures que les parents proches des Prolacertiformes après avoir enlevé caractéristiques du membre postérieur de son analyse, dans une tentative de tester l'idée que ces personnages sont le résultat de évolution convergente entre les ptérosaures et les dinosaures . Cependant, l'analyse ultérieure par Dave Hone et Michael Benton (2007) ne pouvait pas reproduire ce résultat. Hone et Benton trouvé ptérosaures à être étroitement liées aux dinosaures même sans caractères des pattes postérieures. Ils ont également critiqué les études précédentes par David Peters, qui soulève des questions quant à savoir si conclusions sans avoir accès à la meilleure preuve, ce est les fossiles de ptérosaures existantes, peuvent être considérés comme ayant le même poids que des conclusions fondées strictement sur l'interprétation de première main. Peters soutient que DGS est supérieure à à vue et l'interprétation de première main. Hone et Benton a conclu que, bien que pterosauromorphs plus primitives sont nécessaires pour clarifier leurs relations, ptérosaures sont mieux considérées archosaurs et ornithodirans spécifiquement, étant donné les preuves actuelles. Dans l'analyse de Hone et Benton, les ptérosaures sont soit le groupe de soeur de Scleromochlus ou tombent entre elle et Lagosuchus sur l'arbre généalogique ornithodiran. Sterling Nesbitt (2011) a trouvé un appui solide pour un clade composé de Scleromochlus et ptérosaures.
Classification
Classification des ptérosaures a toujours été difficile, car il y avait de nombreuses lacunes dans le registre fossile . Beaucoup de nouvelles découvertes se remplissent ces lacunes et de donner une meilleure image de l'évolution des ptérosaures. Traditionnellement, ils sont organisés en deux sous-ordres:
- Rhamphorhynchoidea (Plieninger, 1901): Un groupe de début, de base ("primitives") ptérosaures, dont beaucoup avait de longues queues et court métacarpiens dans l'aile. Ils sont apparus dans la période du Trias supérieur, et a duré jusqu'à la fin du Jurassique. Est un Rhamphorhynchoidea groupe paraphylétique (depuis les pterodactyloids évolué directement auprès d'eux et non à partir d'un ancêtre commun), donc avec l'utilisation croissante des cladistics Il est tombé en disgrâce dans la plupart littérature technique.
- Pterodactyloidea (Plieninger, 1901): Le plus dérivée («avancé») ptérosaures, avec des queues courtes et longue aile métacarpiens. Ils sont apparus au milieu du Jurassique période, et a duré jusqu'à ce que le Extinction Crétacé-Tertiaire les essuya à la fin des Crétacé .

Liste des familles et superfamilles dans Pterosauria, après Unwin 2006, sauf indication contraire.
- ORDRE Pterosauria (éteint)
- Sous-ordre Rhamphorhynchoidea *
- Famille Anurognathidae
- Famille Campylognathoididae
- Famille Dimorphodontidae
- Famille Rhamphorhynchidae
- Sous-ordre Pterodactyloidea
- Superfamille Ornithocheiridae
- Famille Istiodactylidae
- Famille Nyctosauridae
- Famille Ornithocheiridae
- Famille Pteranodontidae
- Superfamille Ctenochasmatoidea
- Famille Ctenochasmatidae
- Famille Gallodactylidae
- Famille Pterodactylidae
- Superfamille Dsungaripteroidea
- Famille Dsungaripteridae
- Famille Germanodactylidae
- Superfamille Azhdarchoidea
- Famille Azhdarchidae
- Famille Chaoyangopteridae
- Famille Lonchodectidae
- Famille Tapejaridae
- Superfamille Ornithocheiridae
- Sous-ordre Rhamphorhynchoidea *
Les relations précises entre les ptérosaures est toujours en suspens. Cependant, plusieurs études récentes commencent à rendre les choses plus claires. Cladogramme simplifiée après Unwin.
| Pterosauria |
| ||||||||||||||||||||||||||||||
| Pterodactyloidea |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||
Extinction
On pensait autrefois que la concurrence avec les premiers oiseaux espèces aurait abouti à la disparition de la plupart des ptérosaures. À la fin du Crétacé, seules les grandes espèces de ptérosaures sont connus. Les espèces plus petites semblent avoir disparu, leur créneau rempli par les oiseaux. Toutefois, le déclin de ptérosaure (si effectivement présents) semble sans rapport avec la diversité des oiseaux. À la fin de la période du Crétacé, le Crétacé-Paléogène de l'événement d'extinction qui a détruit tous les dinosaures non-aviaires et les dinosaures les plus aviaires ainsi, et de nombreux autres animaux, semblait de prendre également les ptérosaures. Par ailleurs, certains ptérosaures peuvent avoir été spécialisée pour une vie océanique. Par conséquent, lorsque l'événement d'extinction Crétacé-Paléogène sévèrement affecté la vie marine que ces ptérosaures nourris, ils se sont éteints. Cependant, comme forme azhdarchids et istiodactylids marine ne étaient pas dans les habitudes.
Récemment, plusieurs nouveaux taxons de ptérosaures ont été découverts datant du Campanien / Maastrichtien, comme les ornithocheirids Et Piksi " Ornithocheirus ", possible pteranodontids et nyctosaurids, et un tapejarid. Ceci suggère que fin faunes de ptérosaures Crétacé étaient beaucoup plus diversifié qu'on ne le pensait, peut-être même pas avoir diminué de façon significative depuis le début des Crétacé.
Genres bien connus
Des exemples de pterosaur genres suivants:
- Pteranodon était de 1,8 mètres (six pieds) de long, avec une envergure de 7,5 m (25 pi), et a vécu au cours de la fin du Crétacé.
- Pterodactylus avait une envergure de 50 à 75 centimètres (20 à -30 pouces), et a vécu à la fin du Jurassique sur les rives du lac.
- Pterodaustro était un ptérosaure du Crétacé de l'Amérique du Sud avec une envergure d'environ 1,33 mètres et de plus de 500 haut et étroit dents, qui ont sans doute été utilisés dans d'alimentation par filtration, un peu comme modernes flamants . En outre, comme les flamants, l'alimentation de ce ptérosaure peut avoir entraîné l'animal ayant une teinte rose. Ce était la première ptérosaure trouvaille d'Amérique du Sud.
- Quetzalcoatlus avait une envergure de 10 à 11 mètres (33-36 pi), et a été parmi les plus grands animaux volants jamais. Il a vécu pendant la période du Crétacé.
- Ornithocheirus était un grand ptérosaure qui vivait dans le monde entier au cours du Crétacé précoce environ 130 MYA. Les mâles du genre avaient crêtes sur leur facture.
- Hatzegopteryx était le plus grand animal volant, comme on le sait actuellement à la science. Bien qu'aucun fossile complet a été découvert, le peu de fossiles paléontologues ont trouvé la créature donner une envergure estimée à au moins 10 mètres (33 pi)
- Rhamphorhynchus était un ptérosaure jurassienne, avec une palette à la fin de sa queue, qui peut avoir agi pour stabiliser la queue en vol.
Dans la culture populaire

Ptérosaures ont été un aliment de base de la culture populaire aussi longtemps que leurs cousins les dinosaures, mais ils ne sont généralement pas aussi évidente en vedette dans les films, la littérature ou autre art. En outre, tandis que la représentation de dinosaures dans les médias populaires a radicalement changé en réponse aux progrès de la paléontologie, une image essentiellement obsolète de ptérosaures a persisté depuis le milieu du 20e siècle.
Bien que le terme générique "ptérodactyle" est souvent utilisé pour décrire ces créatures, les animaux représentés fréquemment représentent soit Pteranodon ou Rhamphorhynchus, ou un hybride fictif des deux. Les jouets et les dessins animés de nombreux enfants disposent "ptérodactyles» avec Pteranodon -comme crêtes et à long, La queue et des dents de Rhamphorhynchus, une combinaison qui n'a jamais existé dans la nature. Cependant, au moins un type de ptérosaure ne ont au moins crête et des dents, par exemple -comme l'Pteranodon, le Ludodactylus, un nom qui signifie «doigt de jouet" pour sa ressemblance avec les jouets des enfants, vieux inexactes. En outre, certaines représentations de ptérosaures à identifier à tort comme des « oiseaux », quand dans la vie réelle ils volaient les reptiles , et les oiseaux sont en fait les descendants de théropodes dinosaures.
Ptérosaures ont également été utilisés dans la fiction en 1912 roman de Arthur Conan Doyle Le monde perdu, et après 1925 adaptation cinématographique. Ils ont été utilisés dans un certain nombre de films et de programmes de télévision depuis, y compris le film 1933 King Kong, et de 1966 One Million Years BC. Dans ce dernier, animateur Ray Harryhausen a dû ajouter doigts d'ailes de chauve-souris inexactes à ses modèles de mouvement d'arrêt afin de maintenir les membranes de tomber en morceaux, si cette erreur était particulièrement fréquent dans l'art avant même que le film a été réalisé. Ptérosaures étaient essentiellement absents des apparitions au cinéma notables jusqu'en 2001, avec Jurassic Park III . Cependant, paléontologue Dave Hone a noté que, même après les 40 années qui ont suivi, les ptérosaures de ce film avaient pas été mise à jour importante pour refléter la recherche moderne. Parmi les erreurs il a noté que la persistance des années 1960 aux années 2000 étaient les dents, même en espèces sans dent (les Jurassic Park III ptérosaures étaient destinés à être Pteranodon , qui se traduit comme "aile édentée"), le comportement de nidification qui a été connu pour être inexacts en 2001, et ailes de cuir, plutôt que les membranes tendues de la fibre musculaire qui a été effectivement présent et nécessaire pour le vol de ptérosaure.