
Code génétique
À propos de ce écoles sélection Wikipedia
Ce contenu de Wikipedia a été sélectionné par SOS Enfants d'aptitude dans les écoles à travers le monde. Visitez le site Web d'enfants SOS au http://www.soschildren.org/
Le code génétique est un ensemble de règles par lequel l'information codée dans le matériel génétique ( ADN ou des séquences d'ARN) est traduit en protéines ( acides aminés ) par séquences vivant cellules . Plus précisément, le code définit une correspondance entre tri- des séquences de nucleotides appelés codons et acides aminés; chaque triplet de nucleotides dans une séquence d'acide nucléique spécifie un acide aminé unique. Étant donné que la grande majorité des gènes sont encodés avec exactement le même code (voir #RNA codon tableau ), ce code particulier est souvent désigné comme le code génétique canonique ou standard, ou simplement le code génétique, mais en fait il ya de nombreuses variantes de codes; Ainsi, le code génétique canonique ne est pas universelle. Par exemple, chez l'homme, la synthèse des protéines dans les mitochondries se appuie sur un code génétique qui varie à partir du code canonique.
Il est important de savoir que pas toutes les informations génétiques sont stockées en tant que code génétique. L'ADN de tous les organismes de réglementation, contiennent des séquences intergéniques segments, régions chromosomiques structurelles, qui peuvent contribuer grandement à phénotype, mais fonctionner en utilisant un des ensembles distincts de règles qui peuvent ou peuvent ne pas être aussi simple que le paradigme bien définie codon à l'acide aminé qui sous-tend le code génétique.
Décryptage du code génétique
Après la structure de l'ADN a été déchiffrée par James Watson , Francis Crick , Maurice Wilkins et Rosalind Franklin, de sérieux efforts pour comprendre la nature du codage de protéines a commencé. George Gamov postulé qu'un code à trois lettres doit être utilisé pour coder les 20 différents acides aminés utilisés par les cellules vivantes pour coder pour des protéines (parce que 3 est le plus petit n tel que n est égal à 4 au moins 20). Le fait que les codons ne se composent de trois bases de l'ADN a été démontrée dans le premier Crick, Brenner et al. expérience. L'élucidation d'un premier codon a été effectuée par Marshall Nirenberg et Heinrich J. Matthaei en 1961 à la National Institutes of Health. Ils ont utilisé un système acellulaire à traduire une séquence poly-uracile ARN (ou UUUUU ... en termes biochimiques) et découvert que le ils avaient polypeptide synthétisé est composée de seulement l'acide aminé phénylalanine. Ils ainsi déduites de ce poly-phénylalanine que le codon spécifié UUU la phenylalanine amino-acide. L'extension de ce travail, Nirenberg et ses collègues ont pu déterminer la composition nucléotidique de chaque codon. Afin de déterminer l'ordre de la séquence, trinucléotides étaient liés aux ribosomes et radioactivement marqués aminoacyl-ARNt a été utilisé pour déterminer l'acide aminé qui correspond au codon. Le groupe des Nirenberg a pu déterminer les séquences de 54 des 64 codons. Des travaux ultérieurs par Har Gobind Khorana identifié le reste du code, et peu de temps après Robert W. Holley déterminé la structure de ARN de transfert, la molécule d'adaptateur qui facilite la traduction. Ce travail a été basée sur des études antérieures par Severo Ochoa, qui a reçu le prix Nobel en 1959 pour ses travaux sur l'enzymologie de la synthèse de l'ARN. En 1968, Khorana, Holley et Nirenberg a également reçu le Prix Nobel de physiologie ou médecine pour leur travail.
Transfert d'informations via le code génétique
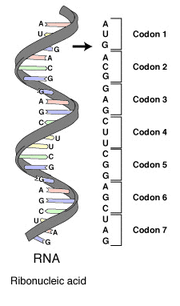
Le génome d'un organisme est inscrit dans l'ADN , ou dans certains virus ARN. La partie du génome qui code pour une protéine ou un ARN est appelé un gène. Les gènes qui codent pour des protéines sont composées d'unités de tri-nucléotides, appelés codons codant chacun pour un acide aminé unique. Chaque sous-unité nucléotidique consiste en un phosphate, sucre de désoxyribose et l'un des quatre azoté bases nucléotidiques. Le bases puriques adénine (A) et guanine (G) sont plus grandes et se composent de deux cycles aromatiques. Le bases pyrimidiques cytosine (C) et thymine (T) sont plus petits et se composent d'un seul cycle aromatique. Dans la configuration en double hélice, deux brins d'ADN sont reliés les uns aux autres par des liaisons hydrogène dans un dispositif connu sous le nom appariement base. Ces obligations font presque toujours entre une base adénine sur un brin et une thymine sur l'autre brin et entre une base cytosine sur un brin et une base de guanine de l'autre. Cela signifie que le nombre de résidus A et T est la même dans une double hélice donné comme le nombre de résidus G et C. Dans l'ARN, la thymine (T) est remplacée par uracile (U), et le désoxyribose est substitué par ribose.
Chaque gène codant pour la protéine est transcrit en une molécule d'ARN de la matrice de polymère est associée, connu sous le nom ARN messager ou ARNm. Ceci à son tour est traduit sur la ribosome dans un acide aminé ou chaîne polypeptide. Le processus de traduction nécessite ARN de transfert spécifiques pour les acides aminés individuels avec les acides aminés attaché de manière covalente à eux, guanosine triphosphate comme source d'énergie, et un certain nombre de facteurs de traduction. ARNt ont anticodons complémentaires aux codons dans l'ARNm et peut être «chargé» de manière covalente avec des acides aminés à son terminal 3 'CCA se termine. ARNt individuels sont facturés avec des acides aminés spécifiques par des enzymes appelées synthétases aminoacyl ARNt qui ont une spécificité élevée pour les deux leurs acides aminés apparentés et ARNt. La grande spécificité de ces enzymes est une raison majeure pour laquelle la fidélité de la traduction des protéines est maintenue.
Il existe des combinaisons 4³ = 64 codon différent possible avec un codon triplet de trois nucleotides. En réalité, les 64 codons de la norme code génétique sont affectés soit à des acides aminés ou des signaux d'arrêt au cours de la traduction. Si, par exemple, une séquence d'ARN, et est considéré comme UUUAAACCC la cadre de lecture commence par le premier U (par convention, 5 'à 3' ), il ya trois codons, à savoir, UUU, AAA et CCC, dont chacun spécifie un acide aminé. Cette séquence d'ARN est traduit en une séquence d'acides aminés, trois acides aminés de long. Une comparaison peut être faite avec l'informatique , où le codon est l'équivalent d'un mot, qui est le "chunk" standard de traitement des données (comme un acide aminé d'une protéine), et un nucléotide pour un bits.
Le code génétique standard est indiqué dans les tableaux suivants. Le tableau 1 montre que l'acide aminé chacun des 64 codons précise. Le tableau 2 montre ce codons spécifient chacun des 20 acides aminés standards impliqués dans la traduction. Ceux-ci sont appelés avant et arrière tables codons, respectivement. Par exemple, le codon AUA de représente l'acide aminé asparagine et UGU et UGC représentent cystéine (désignations à trois lettres standard, Asn et Cys respectivement).
Table ARN codon
| 2e but | |||||
|---|---|---|---|---|---|
| U | C | Un | Sol | ||
| 1er base | U | UUU (Phe / F) Phénylalanine | UCU (Ser / S) Serine | UAU (Tyr / Y) Tyrosine | UGU (Cys / C) Cystéine |
| C | CUU (Leu / L) leucine | CCU (Pro / P) Proline | CAU (Son / H) Histidine | UGT (Arg / R) Arginine | |
| Un | AUU (Ile / I) Isoleucine | ACU (Thr / T) Thréonine | AUA (Asn / N) Asparagine | AGU (Ser / S) Serine | |
| Sol | GUU (Val / V) Valine | GCU (Ala / A) Alanine | GAU (Asp / D) L'acide aspartique | GGU (Gly / G) Glycine | |
| Ala / A | GCU, GCC, GCA, GCG | Leu / L | UUA, UUG, CUU, CUC, AUC, CUG |
|---|---|---|---|
| Arg / R | UGT, CGC, CGA, CGG, AGA, AGG | Lys / K | AAA, AAG |
| Asn / N | AUA, AAC | Met / M | Août |
| Asp / D | GAU, GAC | Phe / F | UUU, UUC |
| Cys / C | UGU, UGC | Pro / P | CCU, CCC, CCA, GCC |
| Gin / Q | CAA, CAG | Ser / S | UCU, UCC, UCA, UCG, AGU, AGC |
| Glu / E | GAA, GAG | Thr / T | ACU, ACC, ACA, ACG |
| Gly / G | GGU, GGC, GGA, GGG | Trp / W | UGG |
| Son / H | CAU, CAC | Tyr / Y | UAU, UAC |
| Ile / I | AUU, AUC, AUA | Val / V | GUU, GUC, GUA, GUG |
| DÉBUT | Août | ARRÊTEZ | UAG, UGA, SAU |
Principales caractéristiques
Le cadre de lecture d'une séquence
Notez qu'un codon est définie par le nucléotide initiale à partir de laquelle commence la traduction. Par exemple, la chaîne GGGAAACCC, se il est lu de la première position, contient les codons GGG, AAA et CCC; et se il est lu à partir de la seconde position, il contient les codons AAC GGA et; se il est lu à partir de la troisième position, GAA et de l'ACC. Codons partiels ont été ignorées dans cet exemple. Chaque séquence peut donc être lu en trois cadres de lecture, dont chacune va produire une séquence d'acides aminés différent (dans l'exemple donné, Gly-Lys-Pro, Gly-Asp, ou Glu-Thr, respectivement). Avec l'ADN double brin, il ya six possibles cadres de lecture, trois dans l'orientation de l'avant sur un brin et trois arrière (sur le brin opposé).
Le cadre réel dans lequel une séquence de protéine est traduite est définie par une codon d'initiation, habituellement le premier codon AUG dans la séquence d'ARNm. Les mutations qui perturbent le cadre de lecture par des insertions ou des deletions d'un non-multiple de trois bases de nucléotides sont connus comme mutations du cadre de lecture. Ces mutations peuvent nuire à la fonction de la protéine résultante, si elle est formée, et sont donc rares dans dans les séquences codant pour des protéines in vivo. Souvent, ces protéines mal formés sont ciblés pour la dégradation protéolytique. En outre, une mutation à décalage cadre est très susceptible de causer un codon d'arrêt à lire qui tronque la création de la protéine (par exemple ). Une raison de la rareté des mutations de cadre étant décalée héritées est que si la protéine en cours de traduction est essentielle à la croissance sous des pressions de sélection de l'organisme doit faire face, l'absence d'une protéine fonctionnelle peut provoquer une létalité avant l'organisme est viable.
START / STOP codons
Traduction commence par une chaîne codon d'initiation (codon d'initiation). Contrairement codons arrêt, le codon seule ne est pas suffisante pour commencer le processus. Séquences proches et facteurs d'initiation sont également tenus de commencer la traduction. Le codon de départ de la plus commune est AUG, qui code pour la methionine, la plupart des chaînes d'acides aminés avec la methionine de départ.
L'arbre codons stop ont été donné des noms: UAG est orange, UGA est opale (parfois aussi appelé la terre d'ombre) et UAA est ocre. "Ambre" a été nommé par les découvreurs Richard Epstein et Charles Steinberg après leur ami Harris Bernstein, dont le nom signifie "orange" en allemand. Les deux autres codons stop ont été nommés «ocre» et «opale» afin de garder le «thème de couleur noms". Codons stop sont aussi appelés codons de terminaison et ils signalent libération du polypeptide naissant du ribosome due à la liaison du libérer des facteurs en l'absence d'ARNt apparenté avec anticodons complémentaire de ces signaux d'arrêt.
La dégénérescence du code génétique,
Le code génétique a la redondance, mais aucune ambiguïté (voir les tableaux codons ci-dessus pour la corrélation complète). Par exemple, bien que codons GAA et GAG fois précisent acide glutamique (redondance), aucun d'entre eux spécifie tout autre acide aminé (pas d'ambiguïté). Les codons codant pour un acide aminé peuvent être différentes l'une quelconque de ses trois positions. Par exemple, l'acide aminé l'acide glutamique est spécifié par GAA et GAG (codons de différence dans la troisième position), les acides aminés leucine est spécifié par UUA, UUG, CUU, CUC, CUA, CUG codons (différence dans le premier ou troisième position), tandis que l'acide aminé sérine est spécifié par UCA, UCG, UCC, UCU, AGU, AGC (différence de la première, deuxième ou troisième position).
Une position d'un codon est dit être un site dégénéré quadruple si ne importe quel nucleotide à cette position spécifie le même acide aminé. Par exemple, la troisième position de la glycine codons (GGA, GGG, GGC, GGU) est un site dégénéré quadruple, parce que toutes les substitutions nucléotidiques sur ce site sont synonymes, ce est à dire qu'ils ne changent pas l'acide aminé. Seule la troisième positions de certains codons peuvent être dégénérée quadruplé. Une position d'un codon est dit être un site dégénérée double si seulement deux des quatre nucleotides possibles à cette position spécifient le même acide aminé. Par exemple, la troisième position de la acide glutamique codons (GAA, GAG) est un site dégénéré double, est donc la première position de la codons leucine (UCA, UCC, CCU, CCC, CCA, GCC). Dans les sites de dégénérés de doubles, les nucléotides équivalentes sont toujours soit deux purines (A / G) ou deux pyrimidines (C / U), des substitutions de sorte que transversional (purine à la pyrimidine ou pyrimidine à purine) dans des sites dégénérés de doubles sont non synonyme. Une position d'un codon est dit être un site non dégénéré si toute mutation à cette position se traduit par la substitution d'acides aminés. Il n'y a qu'un seul site où dégénérée triple changement de trois des quatre nucleotides n'a aucun effet sur l'acide aminé, tout en modifiant les quatrième résultats de nucleotides possibles dans une substitution d'acide aminé. Ce est la troisième position d'un isoleucine codon: AUU, AUC, ou AUA codent tous isoleucine, mais encode août méthionine. Dans le calcul de cette position est souvent considérée comme un site dégénéré double.
Il existe trois acides aminés codés par six codons différents: sérine, la leucine, arginine. Seuls deux acides aminés sont spécifiés par un codon unique; une d'entre elles est l'amino-acide méthionine, spécifiée par le codon AUG, qui précise également le début de la traduction; l'autre est tryptophane, spécifiée par le codon UGG. La dégénérescence du code génétique est ce qui explique l'existence de mutations silencieuses.
La dégénérescence résulte en raison d'un code de triplet désigne 20 acides aminés et un codon d'arrêt. Comme il existe quatre bases, codons triplets sont nécessaires pour produire au moins 21 codes différents. Par exemple, se il y avait deux bases par codon, puis seulement 16 acides aminés pourraient être codé (4² = 16). Parce que au moins 21 codes sont nécessaires, 4³ donne 64 codons possibles, ce qui signifie que certains dégénérescence doit exister.
Ces propriétés du code génétique, il est plus tolérant aux pannes pour des mutations ponctuelles. Par exemple, en théorie, des codons dégénérés quadruplé peuvent tolérer aucune mutation ponctuelle à la troisième position, bien que biais d'usage codon limite dans la pratique dans de nombreux organismes; codons dégénérés de doubles peuvent tolérer une des trois mutations ponctuelles possibles à la troisième position. Depuis mutations de transition (purine à purine ou pyrimidine à des mutations de pyrimidine) sont plus susceptibles que transversion (purine pyrimidine ou vice-versa) mutations, l'équivalence des purines ou des pyrimidines sur les sites double dégénérés ajoute une tolérance aux pannes.

Une conséquence pratique de la redondance est que certaines erreurs dans le code génétique ne provoquent une mutation silencieuse ou une erreur qui ne affecterait pas la protéine parce que le hydrophile ou hydrophobie est maintenu par substitution d'équivalents acides aminés; par exemple, un codon de Nun (où n = ne importe quel nucleotide) tend à coder pour les acides aminés hydrophobes. Rendements NCN résidus d'acides aminés qui sont de petite taille et modérée en hydrothérapie; NAN code taille moyenne résidus hydrophiles; UNN code résidus qui ne sont pas hydrophile.
Même ainsi, les mutations ponctuelles peuvent encore causer des protéines dysfonctionnelles. Par exemple, un muté gène de l'hémoglobine provoque la drépanocytose . Dans l'hémoglobine mutante hydrophile glutamate (Glu) est substitué par le hydrophobe valine (Val), ce qui réduit la solubilité de β-globine. Dans ce cas, les causes de cette mutation l'hémoglobine pour former des polymères linéaires liés par l'interaction hydrophobe entre les groupes valine déformation provoquant la drépanocytose des érythrocytes. La drépanocytose est généralement pas causée par un de novo mutation. Au contraire, il est choisi par le paludisme régions (d'une manière similaire à thalassémie), tel que personnes hétérozygotes ont une certaine résistance à la malaria Plasmodium ( hétérozygote avantage).
Ces codes variables pour les acides aminés sont autorisés à cause de bases modifiées dans la première base de la anticodon de l'ARNt, et la paire formée base est appelée Wobble pairing. Les bases modifiées comprennent inosine et de la paire de bases non-Watson-Crick UG.
Variations au code génétique classique
Bien que de légères variations sur le code standard avaient été prédit plus tôt, aucun n'a été découvert qu'en 1979, lorsque les chercheurs étudiant gènes mitochondriaux humains découverts, ils ont utilisé un code alternatif. Beaucoup de légères variantes ont été découvertes depuis, y compris divers codes mitochondriales alternatifs, ainsi que de petites variantes telles que Mycoplasma traduire le codon UGA comme le tryptophane. Dans les bactéries et archées, GUG et UUG sont codons de départ communs. Cependant, dans de rares cas, certaines protéines spécifiques peuvent utiliser initiation alternatif (départ) des codons pas normalement utilisés par cette espèce.
Dans certaines protéines, des acides aminés non standards sont remplacés par des codons d'arrêt classiques, en fonction des séquences signal associées à l'ARN messager: UGA peut coder pour sélénocystéine et UAG peut coder pour pyrrolysine comme on le verra dans les articles pertinents. Selenocysteine est maintenant considéré comme l'acide aminé 21 et pyrrolysine est considéré comme le 22e. Une description détaillée des variations dans le code génétique peut être trouvé à la NCBI site Web.
Malgré ces différences, tous les codes connus présentent de fortes similitudes avec l'autre, et le mécanisme de codage est la même pour tous les organismes: codons à trois bases, d'ARNt, les ribosomes, la lecture du code dans la même direction et en traduisant le code à trois lettres à la fois en des séquences d'acides aminés.
Théories sur l'origine du code génétique
Malgré les variations qui existent, les codes génétiques utilisées par toutes les formes connues de la vie sur Terre sont très similaires. Comme il existe de nombreux codes génétiques possibles qui sont censées être utilitaire similaire à celui utilisé par la vie de la Terre, la théorie de l'évolution suggère que le code génétique a été établie très tôt dans l'histoire de la vie, avec une analyse phylogénétique des ARN de transfert suggère que ARNt molécules évolué avant que l'ensemble présente des aminoacyl-ARNt synthétases.
Le code génétique ne est pas une assignation aléatoire de codons d'acides aminés. Par exemple, les acides aminés qui partagent la même voie de biosynthèse ont tendance à avoir la même première base de leurs codons, et des acides aminés ayant des propriétés physiques similaires ont tendance à avoir des codons similaires.
Il ya trois thèmes qui traversent les nombreuses théories qui cherchent à expliquer l'évolution du code génétique (et donc l'origine de ces modèles). Une est illustrée par la récente aptamères expériences qui montrent que certains acides aminés ont une affinité chimique sélective de la base triplets qui codent pour eux. Ceci suggère que le mécanisme de translation de courant, complexe impliquant ARNt et les enzymes peuvent être associés un développement ultérieur, et que l'origine des séquences de protéines ont été directement sur matrice de séquences de bases. Une autre est que le code génétique classique que nous voyons aujourd'hui est passé de, un code plus simple plus tôt par un processus d '«expansion biosynthétique". Ici, l'idée est que la vie primordiale »découvert« nouveaux acides aminés (par exemple, les sous-produits du métabolisme) et plus tard, de retour incorporé certains d'entre eux dans la machine de codage génétique. Bien que beaucoup de preuves circonstancielles a été trouvé pour suggérer que moins de différents acides aminés ont été utilisés dans le passé qu'aujourd'hui, des hypothèses précises et détaillées sur exactement quels acides aminés entré le code exactement de quel ordre se est révélée beaucoup plus controversée. Une troisième théorie est que la sélection naturelle a conduit à des affectations codons du code génétique qui minimisent les effets de mutations.